AlphaFold 3 vs. Molecular Docking: A Critical Evaluation for Pose Prediction in Drug Discovery
This article provides a comprehensive evaluation of AlphaFold 3's capabilities for protein-ligand pose prediction against established molecular docking methods.
AlphaFold 3 vs. Molecular Docking: A Critical Evaluation for Pose Prediction in Drug Discovery
Abstract
This article provides a comprehensive evaluation of AlphaFold 3's capabilities for protein-ligand pose prediction against established molecular docking methods. Aimed at researchers and drug development professionals, it explores the foundational principles of co-folding and docking, analyzes performance benchmarks across diverse biological targets, and addresses critical limitations such as physical realism and generalization. The review offers practical guidance for troubleshooting predictions, leveraging confidence metrics, and integrating these tools into robust research workflows. By synthesizing recent validation studies and comparative analyses, this resource aims to equip scientists with the knowledge to effectively and critically apply these powerful technologies in structural biology and drug design.
The New Paradigm: Understanding Co-Folding and Traditional Docking
The accurate prediction of how small molecules interact with protein targets is a cornerstone of modern drug discovery. For decades, the dominant computational approach has been search-and-score molecular docking, a method that relies on physics-inspired scoring functions to evaluate millions of potential ligand poses. However, the recent advent of deep learning co-folding models, exemplified by AlphaFold 3 (AF3), promises a paradigm shift. This guide provides an objective comparison of these two methodologies for pose prediction research, framing them within a broader thesis on their respective capabilities, limitations, and optimal applications. By synthesizing findings from recent independent benchmarks and original research, we aim to equip researchers with the data needed to select the appropriate tool for their specific scientific question.
Core Methodological Principles
Search-and-Score Molecular Docking
Traditional molecular docking operates on a search-and-score framework [1]. It involves computationally sampling a vast space of possible ligand conformations and orientations (the "search") within a defined protein binding pocket. Each candidate pose is then evaluated using a scoring function—an algorithmic approximation of the binding affinity—to identify the most probable binding mode [1] [2]. These scoring functions can be physics-based (considering van der Waals forces, electrostatics, etc.), empirical (parameterized against experimental binding data), or knowledge-based [2]. A significant limitation of most traditional methods is their treatment of the protein receptor as a rigid body, which fails to capture the induced fit conformational changes that often occur upon ligand binding [1].
Co-Folding with AlphaFold 3
AlphaFold 3 represents a fundamentally different approach. It is a deep learning model that uses a diffusion-based architecture to predict the joint 3D structure of a biomolecular complex from scratch, using only the protein sequence and the ligand's SMILES string as input [3]. Instead of searching and scoring, AF3 "co-folds" the molecules into their bound configuration. Its key innovation is the replacement of AlphaFold 2's structure module with a diffusion module that predicts raw atom coordinates directly, eliminating the need for complex, molecule-specific representations and losses [3]. This allows AF3 to model complexes of proteins, nucleic acids, ions, and small molecules within a single, unified framework.
Visual Comparison of Workflows
The diagram below illustrates the fundamental differences in the operational workflows between a traditional docking pipeline and the AlphaFold 3 co-folding process.
Performance Benchmarking and Experimental Data
Independent studies have rigorously evaluated the performance of AF3 against established docking tools. The following tables summarize key quantitative findings from these benchmarks.
Table 1: Overall Protein-Ligand Docking Accuracy on the PoseBusters Benchmark [3] [4] [5]
| Method | Input Requirements | Success Rate (PB-Valid & <2Å RMSD) | Notes |
|---|---|---|---|
| AlphaFold 3 (Blind) | Protein Sequence, Ligand SMILES | ~76% | No structural input; true blind docking |
| AlphaFold 3 (Pocket-Specified) | Protein Sequence, Ligand SMILES, Pocket Residues | ~93% | Informed of binding site location [3] |
| Strong Baseline (Vina + Gnina) | Protein Structure, Ligand SMILES | ~80% | Uses experimental receptor structure [4] |
| Original Vina Baseline | Protein Structure, Ligand SMILES | ~61% | As reported in AF3 paper [3] [4] |
| DiffDock | Protein Structure, Ligand SMILES | ~38% | Previous leading deep learning docking method [6] |
Table 2: Performance on Specific Docking Challenges and Complex Types [6] [7] [5]
| Task / Complex Type | Representative Method | Performance Metric | Result / Limitation |
|---|---|---|---|
| Antibody-Antigen Docking | AlphaFold 3 (single seed) | High-Accuracy Success Rate (DockQ ≥0.8) | 10.2% (Antibody), 13.3% (Nanobody) [7] |
| AlphaFold 3 (1,000 seeds) | Overall Docking Success Rate | ~60% [7] | |
| Binding Site Mutagenesis | Co-folding models (AF3, RFAA, etc.) | Robustness to non-physical binding site mutations | Poor; models place ligands in mutated sites despite loss of interactions [6] |
| Covalent Ligand Prediction | AlphaFold 3 | AUC for classifying binders vs. decoys | 98.3% [5] |
| Unseen vs. Common Ligands | AlphaFold 3 (Blind) | Success Rate on common natural ligands | Excels (e.g., nucleotides) [4] |
| Strong Baseline | Success Rate on drug-like molecules (excl. common naturals) | Outperforms blind AF3 by 8.5% [4] |
Key Experimental Protocols in Benchmarking
To critically assess the results, it is essential to understand the methodologies behind these benchmarks:
- PoseBusters Benchmark [3] [4]: This test set comprises 428 protein-ligand crystal structures released after AF3's training data cutoff. The primary success metric is the percentage of predictions where the ligand's pocket-aligned Root Mean Square Deviation (RMSD) is less than 2.0 Å and the pose is free of stereochemical violations and severe clashes (deemed "PB-valid").
- Adversarial Physical Challenge [6]: This protocol tests the model's understanding of physical principles rather than its pattern recognition. Researchers selected a known complex (e.g., ATP-bound CDK2) and mutated all binding site residues to glycine (removing side-chain interactions) or phenylalanine (sterically occluding the pocket). A physically intuitive model should predict the ligand is displaced, whereas an overfitted one may still place it in the original site.
- Antibody-Antigen Docking Benchmark [7]: This involves a curated, redundancy-filtered set of antibody and nanobody complexes. Performance is evaluated using the DockQ score, which integrates interface and ligand RMSD metrics into a single value, with DockQ ≥ 0.8 indicating "high-accuracy" predictions as per CAPRI standards.
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Software Tools and Resources for Pose Prediction Research
| Tool / Resource | Type | Primary Function | Access |
|---|---|---|---|
| AlphaFold Server | Web Server | Provides free access to AlphaFold 3 for non-commercial research. | Public Web Interface |
| Gnina | Software | A deep learning-based scoring function for rescoring and selecting docking poses from tools like Vina [4]. | Open-Source |
| ABCFold | Software Toolbox | Simplifies the execution and comparison of AF3, Boltz-1, and Chai-1 by standardizing inputs and outputs [5]. | Open-Source |
| PoseBusters | Python Package | Validates and checks the physical realism and quality of predicted molecular poses against experimental structures [4]. | Open-Source |
| Boltz-1 / Boltz-2 | Software | AF3-like models that introduce features like user-defined pocket conditioning and binding affinity prediction [5]. | Varies |
| Chai-1 | Software | An AF3-like multi-modal foundation model that can be prompted with experimental constraints [5]. | Python Package |
| FeatureDock | Software | A transformer-based docking method that predicts ligand probability density envelopes, useful for pose prediction and scoring [2]. | Open-Source |
Comparative Analysis: Strengths and Limitations
Advantages of AlphaFold 3 Co-Folding
- Elimination of the Protein Structure Requirement: AF3's most significant advantage is its ability to perform true blind docking using only a protein sequence, making it invaluable for targets with no experimentally solved structures [3] [4].
- High Accuracy in Favorable Conditions: When the binding site is known and provided as input, or for ligands highly represented in its training data (e.g., common natural ligands), AF3's accuracy is exceptional, often surpassing traditional methods [3] [4] [5].
- Unified Framework: AF3 can model a vast array of biomolecular interactions—proteins, nucleic acids, ions, and small molecules—within a single model, offering remarkable versatility [3] [8].
Limitations and Concerns of AlphaFold 3
- Questionable Physical Understanding: Adversarial challenges reveal that AF3 and similar co-folding models can fail to learn underlying physics. They sometimes prioritize memorizing common binding motifs over modeling the specific chemical interactions of a given system, leading to physically implausible predictions when binding sites are mutated [6].
- Performance on Drug-like Molecules: While AF3 excels on common natural ligands, its performance advantage narrows or disappears on more "drug-like" molecules, particularly those with halogens or other features less common in the PDB. In one assessment, a strong traditional baseline outperformed blind AF3 on this subset of molecules [4].
- Sampling and Resource Intensity: Achieving top performance, especially for challenging targets like antibody-antigen complexes, can require generating hundreds or thousands of seeds (predictions), which is computationally expensive and, on the public server, subject to job limits [7] [5].
- Bias Towards Training Data: AF3 has been shown to exhibit biases, such as a tendency to predict active-state conformations of GPCRs regardless of the ligand's pharmacological type (agonist/antagonist) [5].
Persistent Strengths of Traditional Docking
- Strong Baselines are Competitive: When a high-quality experimental protein structure is available, robust traditional pipelines (e.g., using conformational ensembles and neural network rescoring with Gnina) can achieve accuracy comparable to or even exceeding blind AF3, and closely approaching pocket-specified AF3 performance [4].
- Explicit Handling of Physics: Traditional methods, while approximate, are built upon physical principles and force fields. This provides a more transparent, interpretable foundation for pose evaluation, even if the scoring functions are imperfect [1] [2].
- Computational Efficiency for Virtual Screening: Well-optimized docking programs are generally faster and more resource-efficient for screening ultra-large libraries of compounds than running multiple AF3 seeds per ligand [1] [4].
Integrated Workflow and Future Directions
The evidence suggests that AF3 and traditional docking are not simply replacements for one another but can be complementary tools. A synergistic workflow is emerging:
- Blind Site Identification with AF3: For targets with unknown or uncertain binding sites, use AF3 for blind prediction to identify potential binding pockets.
- Pose Generation and Refinement: Use the AF3-predicted structure or an available experimental structure with traditional docking (enhanced with conformational sampling and ML rescoring) to generate and rank poses for a library of ligands.
- Cross-Validation and Analysis: Employ tools like PoseBusters to validate the physical realism of the final predictions from both methods.
Future developments will likely focus on improving the physical robustness of deep learning models [6], integrating protein flexibility more effectively [1], and creating more seamless hybrid workflows that leverage the unique strengths of both co-folding and search-and-score paradigms.
The field of computational structural biology has undergone a revolutionary transformation with the introduction of DeepMind's AlphaFold models. While AlphaFold 2 (AF2) demonstrated unprecedented accuracy in protein structure prediction through its innovative Evoformer architecture, AlphaFold 3 (AF3) represents a fundamental paradigm shift by replacing the structure module with a diffusion-based approach and extending capabilities beyond proteins to a wide range of biomolecules [9] [3]. This architectural evolution enables researchers to predict the joint structure of complexes comprising proteins, nucleic acids, small molecules, ions, and modified residues within a single unified deep-learning framework [3]. The implications for drug discovery are profound, as AF3 demonstrates at least 50% better accuracy than existing methods for protein-molecule interactions, with accuracy for specific cases like protein-ligand binding reportedly doubling [10]. This guide provides a comprehensive technical comparison of these architectures, their performance benchmarks, and practical implications for pose prediction research.
Architectural Comparison: Evoformer vs. Diffusion
AlphaFold 2's Evoformer Architecture
AlphaFold 2's architecture centers on the Evoformer module, a specialized transformer network that jointly processes both the multiple sequence alignment (MSA) representation and the pair representation [9] [11]. The system operates through several key components:
- Input Embeddings: AF2 utilized 23 tokens representing the 20 standard amino acids, plus tokens for unknown amino acids, gaps, and masked MSA positions [9]
- Evoformer Stack: The core innovation that operates over both MSA and residue pairs through attention mechanisms and triangular multiplicative updates [12]
- Structure Module: This component generated atomic coordinates using a frame-based representation centered on Cα atoms, with side chains parameterized by χ-angles [13]. It required carefully tuned stereochemical violation penalties during training to enforce chemical plausibility [3]
The system was trained with specialized losses to maintain physical realism and achieved remarkable accuracy by leveraging evolutionary information from MSAs [11].
AlphaFold 3's Diffusion Architecture
AlphaFold 3 introduces substantial modifications to accommodate general biomolecular modeling:
- Expanded Input Tokens: Incorporates tokens for DNA, RNA, and general molecules (represented by single heavy atoms) [9]
- Simplified Trunk Architecture: Replaces the Evoformer with a Pairformer that processes only single and pair representations, removing the MSA representation from core processing [3] [13]
- Diffusion Module: The most significant change—replaces the structure module with a diffusion approach that operates directly on raw atom coordinates without rotational frames or equivariant processing [3]
The diffusion approach employs a relatively standard diffusion model trained to receive "noised" atomic coordinates and predict the true coordinates [3]. This method requires the network to learn protein structure at multiple length scales, with denoising at small noise levels emphasizing local stereochemistry and high noise levels emphasizing large-scale structure [3]. A notable advantage is the elimination of both torsion-based parameterizations and violation losses while handling the full complexity of general ligands [3].
Table 1: Core Architectural Components Comparison
| Component | AlphaFold 2 | AlphaFold 3 |
|---|---|---|
| Core Architecture | Evoformer (processes MSA + pair representations) | Pairformer (processes only single + pair representations) |
| Structure Generation | Structure module with frame-based representation | Diffusion model operating on raw atom coordinates |
| Molecular Representation | Protein-specific (Cα frames with χ-angles) | Universal atomic-level representation |
| Input Scope | Proteins only | Proteins, nucleic acids, ligands, ions, modifications |
| Spatial Inductive Bias | Equivariant transformations | Minimal spatial bias with position embedding |
Performance Benchmarks Across Biomolecular Complexes
Protein-Ligand Interactions
AF3 demonstrates substantial improvements in protein-ligand docking accuracy compared to both traditional docking tools and specialized machine learning approaches:
Table 2: Protein-Ligand Docking Performance on PoseBusters Benchmark
| Method | Category | Accuracy (Ligand RMSD < 2Å) |
|---|---|---|
| AlphaFold 3 | Blind prediction | Significantly outperforms all methods |
| Vina | Traditional docking (uses structural inputs) | Substantially lower than AF3 |
| RoseTTAFold All-Atom | Blind prediction | Much lower than AF3 |
| AF3 (Early Training Cutoff) | Blind prediction | ~40-80% depending on modification type [9] |
On the PoseBusters benchmark (428 protein-ligand structures from PDB released in 2021 or later), AF3 "greatly outperforms classical docking tools such as state-of-the-art Vina" even without using structural inputs that traditional docking methods typically require [3]. The accuracy varies by modification type, with approximately 80% accuracy for bonded ligands and 40% for RNA-modified residues, though statistical error is relatively high due to limited dataset sizes [9].
Protein-Protein and Antibody-Antigen Complexes
AF3 shows notable improvements in protein-protein interactions, with particularly significant gains in antibody-antigen modeling:
Table 3: Antibody-Antigen Docking Performance
| Method | High-Accuracy Success Rate (DockQ ≥0.8) | Overall Success Rate (DockQ ≥0.23) |
|---|---|---|
| AlphaFold 3 (single seed) | 10.2% (antibodies), 13.3% (nanobodies) | 34.7% (antibodies), 31.6% (nanobodies) |
| AlphaFold-Multimer v2.3 | 2.4% | 23.4% |
| AlphaFold 3 (1,000 seeds) | ~60% (as reported by DeepMind) | Not specified |
| Boltz-1 | 4.1% (antibodies), 5.0% (nanobodies) | 20.4% (antibodies), 23.3% (nanobodies) |
Despite these improvements, a recent evaluation noted that AF3 has a 65% failure rate for antibody and nanobody docking with single seed sampling, "demonstrating a need to further improve antibody modeling tools" [7]. The same study found that while AF3 achieves better direct prediction-experiment comparisons, after molecular dynamics simulation relaxation, "the quality of structural ensembles sampled drops severely," potentially due to "instability of the predicted intermolecular packing" [14].
Protein-Nucleic Acid Complexes
AF3 achieves higher accuracy in predicting protein-nucleic acid complexes and RNA structures compared to specialized state-of-the-art tools like RoseTTAFold2NA and AIchemyRNA (the best AI submission of CASP15) on CASP15 examples and a PDB protein-nucleic acid dataset [9]. However, on the CASP15 benchmark, the best human-expert-aided AIchemyRNA2 performed slightly better than AF3 [9].
Experimental Protocols and Methodologies
Standard Benchmarking Protocols
Key experiments evaluating AF3's performance follow standardized protocols:
PoseBusters Protein-Ligand Benchmark:
- Dataset: 428 protein-ligand structures from PDB released in 2021 or later [3]
- Metric: Percentage of protein-ligand pairs with pocket-aligned ligand root mean squared deviation (RMSD) < 2Å
- Comparison Groups: True blind docking methods (similar inputs to AF3) and traditional docking tools that use structural information from solved complexes [3]
Antibody-Antigen Docking Evaluation:
- Dataset: Curated benchmark sets of bound and unbound antibodies/nanobodies from SAbDab, filtered by AF3's training set cutoff with quality and redundancy filtering [7]
- Metrics: DockQ score categorizing predictions as incorrect (DockQ<0.23), acceptable (0.23-0.49), medium (0.49-0.8), or high accuracy (≥0.8) [7]
- Sampling: Typically 1-3 seeds for standard evaluation, with DeepMind reporting results with up to 1,000 seeds [7]
Cross-Docking and Apo-Docking Challenges:
- Cross-docking: Ligands docked to alternative receptor conformations from different ligand complexes [1]
- Apo-docking: Uses unbound (apo) receptor structures from crystal structures or computational predictions [1]
- Significance: These represent realistic drug discovery scenarios where proteins undergo conformational changes upon binding [1]
Critical Analysis of Experimental Findings
While benchmarks show impressive performance, several studies note important limitations:
- Physical Realism: AF3 predictions show "major inconsistencies/deviations from experiment in the compactness of the complex, the intermolecular directional polar interactions (>2 hydrogen bonds are incorrectly predicted) and interfacial contacts" [14]
- Sampling Limitations: Single seed sampling yields limited success (34.7% for antibodies), requiring extensive sampling (1,000 seeds) to achieve 60% success rates [7]
- Generalization Challenges: Performance drops when docking to apo structures or handling significant conformational changes [1]
Practical Implementation and Research Applications
The Scientist's Toolkit: Essential Research Reagents
Table 4: Key Research Resources for AlphaFold-Based Studies
| Resource | Type | Function and Application |
|---|---|---|
| AlphaFold Server | Web Platform | Free academic access for non-commercial prediction of complexes [10] |
| PDBbind | Database | Curated protein-ligand complexes for training and benchmarking [1] |
| PoseBusters | Benchmarking Suite | Validates structural plausibility and assesses prediction quality [3] |
| SAbDab | Database | Structural antibody database for antibody-specific benchmarks [7] |
| UniProt | Database | Protein sequences and annotations for MSA construction [9] |
Research Workflow Integration
For pose prediction research, integrating AF3 requires careful consideration:
Input Preparation:
- Proteins: Amino acid sequences
- Nucleic acids: Nucleotide sequences
- Small molecules: SMILES strings [3]
- Modified residues: Specification of modifications (phosphorylation, glycosylation, etc.)
Quality Evaluation:
- Confidence metrics: pLDDT (per-residue confidence), pTM (predicted TM-score), ipTM (interface pTM) [3] [15]
- Structural validation: Clash scores, bond geometry, steric interactions [1]
Hybrid Approaches:
- Use AF3 for initial pose generation followed by physics-based refinement [1] [14]
- Combine with molecular dynamics for ensemble generation [14]
- Integrate with binding affinity prediction tools [1]
Architectural Workflow Visualization
Architecture Evolution from AF2 to AF3 - This diagram illustrates the fundamental architectural shift from AlphaFold 2's Evoformer-based processing to AlphaFold 3's Pairformer and diffusion-based approach, highlighting the expansion from protein-only to general biomolecular modeling.
The architectural evolution from AlphaFold 2's Evoformer to AlphaFold 3's diffusion model represents a significant advancement in biomolecular structure prediction. The key advantages of AF3 include:
- Broader Biomolecular Scope: Capacity to model nearly all molecular types in the PDB within a unified framework [3]
- Improved Accuracy: Substantially higher accuracy across most interaction categories compared to specialized tools [3]
- Physical Plausibility: Elimination of explicit stereochemical constraints while maintaining physically realistic structures [3]
However, important limitations remain for researchers considering AF3 for pose prediction:
- Sampling Intensity: High accuracy often requires extensive sampling (many seeds) [7]
- Dynamic Processes: Provides structural snapshots rather than dynamic conformational changes [10]
- Commercial Restrictions: Limited availability for commercial drug discovery [10]
- RNA Challenges: Mixed performance on RNA structure prediction [10]
For molecular docking research, AF3 represents a powerful tool that excels at generating accurate initial poses but benefits from integration with physics-based refinement methods and experimental validation. The architectural shift from domain-specific parameterizations to a universal diffusion approach suggests a promising direction for future biomolecular modeling, though careful validation remains essential, particularly for therapeutic applications.
In the field of computational drug discovery, the prediction of protein-ligand binding poses represents a fundamental challenge with significant implications for pharmaceutical development. Researchers increasingly rely on two divergent methodological paradigms: deep learning-based co-folding models like AlphaFold 3 and physics-based molecular docking approaches. However, a critical but often overlooked factor unites these seemingly disparate methodologies—their shared dependence on the quality and composition of the training data they utilize. At the center of this data ecosystem sits PDBbind, a curated database of protein-ligand complexes and their binding affinities that has become the de facto standard for training and validating predictive models. While this resource has been invaluable to the community, evidence suggests that the database's structural artifacts, statistical anomalies, and organization may inadvertently encourage models to memorize specific data patterns rather than learn the underlying physics of molecular interactions. This review examines how PDBbind's characteristics shape the learning behaviors of both deep learning and traditional docking approaches, with profound implications for their real-world performance in pose prediction research.
PDBbind Under the Microscope: Structural and Statistical Challenges
Documented Data Quality Issues
The PDBbind database, while instrumental in advancing computational drug discovery, suffers from several documented quality concerns that may compromise the accuracy and generalizability of models trained upon it. A recent analysis of PDBbind v2020 revealed several common structural artifacts affecting both proteins and ligands, including incorrect bond orders, unreasonable protonation states, and missing atoms in protein chains [16]. Perhaps more critically, the database contains severe steric clashes between protein and ligand heavy atoms at distances closer than 2 Ångstroms, which represent physically implausible non-covalent interactions that can misdirect the learning process of predictive algorithms [16].
The curation process itself presents additional challenges. The PDBbind data processing procedure is neither open-sourced nor automated, potentially relying on manual intervention that can introduce inconsistencies across different entries [16]. This lack of transparency and standardization complicates efforts to reproduce results or identify systematic errors in the dataset.
The Data Leakage Problem
Perhaps the most significant challenge for rigorous model evaluation is the issue of data leakage within PDBbind's standard data splits. The general, refined, and core datasets are cross-contaminated with proteins and ligands exhibiting high similarity [17]. This contamination artificially inflates performance metrics when models are tested on protein-ligand complexes that closely resemble those in their training data, creating a false confidence in their predictive capabilities for truly novel targets [17].
The conventional random splitting of PDBbind into training and test sets fails to account for similarities in protein sequences and ligand chemical structures, allowing models to perform well through a form of "short-term memorization" of analogous patterns rather than genuinely learning the principles of molecular recognition [17]. This problem persists even in time-based splits, as new drugs frequently target established protein families, and existing compounds are often tested against new protein targets [17].
Experimental Evidence: Case Studies in Data Dependency
HiQBind-WF: A Diagnostic Workflow
In response to PDBbind's structural issues, researchers developed HiQBind-WF, a semi-automated workflow that diagnoses and corrects common artifacts in protein-ligand complexes [16]. The workflow employs multiple filtering modules to create higher-quality datasets:
- Covalent Binder Filter: Excludes ligands covalently bound to proteins, focusing specifically on non-covalent interactions [16]
- Rare Element Filter: Removes ligands containing elements beyond H, C, N, O, F, P, S, Cl, Br, I to reduce data sparsity [16]
- Small Ligand Filter: Excludes ligands with fewer than 4 heavy atoms, eliminating small inorganic binders beyond typical drug discovery scope [16]
- Steric Clashes Filter: Discards structures with protein-ligand heavy atom pairs closer than 2 Å, removing physically impossible interactions [16]
When applied to PDBbind v2020, this workflow demonstrated significant corrections to structural imperfections, suggesting that models trained on the original dataset may learn from—and potentially memorize—erroneous structural features [16].
LP-PDBBind: Addressing Data Leakage
The Leak Proof PDBBind (LP-PDBBind) dataset represents a systematic effort to reorganize PDBbind to control for data leakage [17]. This approach implements similarity control on both proteins and ligands across training, validation, and test sets, ensuring that models are evaluated on truly novel complexes rather than variations of familiar patterns [17]. The cleaning process also removes covalent complexes and resolves energy unit inconsistencies, creating a more reliable benchmark for assessing model generalizability.
When popular scoring functions including AutoDock Vina, RF-Score, IGN, and DeepDTA were retrained on LP-PDBBind and evaluated on the independent BDB2020+ dataset, they demonstrated significantly better generalization compared to models trained on standard PDBbind splits [17]. This performance gap reveals the extent to which conventional benchmarking approaches have overestimated model capabilities due to data leakage.
Table 1: Performance Comparison of Models Trained on Standard vs. Leak-Proof PDBBind
| Scoring Function | Training Dataset | Performance on PDBBind Core | Performance on BDB2020+ | Generalization Gap |
|---|---|---|---|---|
| AutoDock Vina | Standard PDBBind | High | Moderate | Significant |
| AutoDock Vina | LP-PDBBind | Moderate | High | Small |
| IGN | Standard PDBBind | Very High | Moderate | Large |
| IGN | LP-PDBBind | High | High | Small |
| RF-Score | Standard PDBBind | High | Low | Very Large |
| RF-Score | LP-PDBBind | Moderate | Moderate | Small |
BindingNet: Expanding Data Diversity
The limitations of PDBbind's size and diversity have prompted efforts to create expanded datasets like BindingNet v2, which comprises 689,796 modeled protein-ligand binding complexes across 1,794 protein targets [18]. This represents a substantial expansion beyond PDBbind's approximately 19,500 complexes, offering greater chemical and structural diversity for training [16] [18].
When the Uni-Mol model was trained exclusively on PDBbind, it achieved only a 38.55% success rate (ligand RMSD < 2Å) for novel ligands with low similarity (Tanimoto coefficient < 0.3) to training examples [18]. However, when trained with progressively larger subsets of BindingNet v2, its performance improved dramatically to 64.25%, demonstrating how limited data diversity forces models to interpolate rather than generalize [18]. With the addition of physics-based refinement, the success rate further increased to 74.07% while passing PoseBusters validity checks [18].
Table 2: Performance Improvement with Expanded Training Data (Uni-Mol Model)
| Training Dataset | Success Rate (Novel Ligands) | Passes PoseBusters Validity | Generalization Ability |
|---|---|---|---|
| PDBbind only | 38.55% | No | Low |
| PDBbind + BindingNet v2 (small) | 54.21% | Partial | Moderate |
| PDBbind + BindingNet v2 (medium) | 57.71% | Partial | Moderate |
| PDBbind + BindingNet v2 (full) | 64.25% | Yes | High |
| PDBbind + BindingNet v2 + Physics Refinement | 74.07% | Yes | Very High |
AlphaFold 3 vs. Molecular Docking: A Data-Divide Perspective
AlphaFold 3's Co-Folding Approach
AlphaFold 3 represents a significant advancement in structure prediction through its unified deep-learning framework that jointly models complete molecular complexes [3]. By employing a diffusion-based architecture, AF3 predicts the raw atom coordinates of complexes containing proteins, nucleic acids, small molecules, ions, and modified residues without relying on rotational frames or torsion angle representations [3] [19]. This approach demonstrates substantially improved accuracy over previous specialized tools, achieving approximately 81% accuracy on blind docking benchmarks compared to 38% for DiffDock and 60% for AutoDock Vina when the binding site is provided [6] [3].
However, AF3's performance appears contingent on patterns in its training data. When subjected to adversarial examples based on physical principles, the model demonstrates notable discrepancies in protein-ligand structural predictions [6]. In binding site mutagenesis challenges where all contact residues were replaced with glycine or phenylalanine, AF3 continued to predict similar binding modes despite the removal of crucial interactions, suggesting potential overfitting to specific data features in its training corpus [6].
Molecular Docking's Physical Priors
Traditional molecular docking approaches like AutoDock Vina employ physics-inspired scoring functions that explicitly model intermolecular interactions such van der Waals forces, hydrogen bonding, and electrostatic complementarity [20] [17]. While these methods generally show lower pose prediction accuracy than AF3 on standard benchmarks, they maintain more consistent performance across structural variations because their physical priors provide a form of regularization against memorization [6] [17].
The performance gap between these approaches narrows significantly when evaluated under rigorous data splitting protocols. Docking methods show less performance degradation than deep learning models when moving from standard PDBbind benchmarks to truly independent test sets, suggesting that their physical basis provides better generalization to novel targets [17].
The Hybrid Future
Emerging research suggests that the most promising path forward may integrate both approaches. One study combined deep learning pre-screening with molecular docking validation to identify potential SARS-CoV-2 main protease inhibitors [20]. This hybrid framework leveraged the pattern recognition strength of deep learning with the physical plausibility guarantees of docking, ultimately identifying Enasidenib as a promising candidate that met all selection criteria [20].
Similarly, the integration of physics-based refinement with deep learning pose prediction in the BindingNet study increased success rates by nearly 10 percentage points while ensuring physical validity [18]. These approaches acknowledge that while deep learning can identify promising regions of chemical space, physical simulation remains essential for verifying mechanistic plausibility.
Experimental Protocols for Rigorous Evaluation
Binding Site Mutagenesis Protocol
To assess whether models learn physical principles or memorize training examples, researchers have developed a binding site mutagenesis protocol [6]:
- Select a protein-ligand complex with a known crystal structure (e.g., ATP binding to CDK2)
- Systematically mutate binding site residues through increasingly drastic perturbations:
- Replace all contact residues with glycine to remove side-chain interactions
- Replace all contact residues with phenylalanine to sterically occlude the pocket
- Replace each residue with chemically dissimilar alternatives to alter shape and properties
- Predict the ligand binding pose for each mutated complex using the model being evaluated
- Measure the RMSD between predicted poses and the original crystal structure reference
- Identify steric clashes and physically implausible interactions in the predictions
Models that understand physics should predict ligand displacement when favorable interactions are removed, while models that memorize training data will continue predicting similar binding modes despite unfavorable conditions [6].
Time-Split Cross-Validation Protocol
To properly evaluate generalization to novel targets, researchers recommend a time-split cross-validation approach [17]:
- Collect protein-ligand complexes from PDBbind and timestamp them by their deposition date
- Train models exclusively on complexes deposited before a specific cutoff date (e.g., 2019)
- Evaluate model performance on complexes deposited after the cutoff date
- Calculate similarity metrics between training and test complexes using:
- Protein sequence alignment scores
- Ligand chemical similarity (Tanimoto coefficients)
- Binding site structural similarity
- Stratify performance results by similarity levels to identify performance cliffs
This protocol more closely mimics real-world drug discovery scenarios where models are applied to newly determined targets rather than variations of familiar ones [17].
Visualization of Methodologies and Relationships
The diagram below illustrates the core methodologies and their relationship to training data in protein-ligand pose prediction.
Table 3: Key Research Reagents and Computational Resources
| Resource Name | Type | Primary Function | Key Features/Benefits |
|---|---|---|---|
| PDBbind | Database | Curated protein-ligand complexes with binding affinities | ~19,500 complexes; standard benchmark; includes "general", "refined", and "core" subsets [16] |
| HiQBind-WF | Computational workflow | Corrects structural artifacts in protein-ligand complexes | Fixes bond orders, protonation states, missing atoms; removes steric clashes [16] |
| LP-PDBbind | Reorganized dataset | Data splits controlling for protein/ligand similarity | Prevents data leakage; enables true generalization assessment [17] |
| BindingNet v2 | Expanded dataset | Modeled protein-ligand complexes | 689,796 complexes across 1,794 targets; expands chemical diversity [18] |
| AlphaFold Server | Web service | Predicts biomolecular complex structures | Free academic access; handles proteins, nucleic acids, small molecules [10] |
| AutoDock Vina | Docking software | Predicts ligand binding modes and affinities | Physics-inspired scoring; widely used; open source [20] [17] |
| PoseBusters | Validation suite | Checks physical plausibility of predicted complexes | Detects steric clashes, bond length violations, other artifacts [3] [18] |
| BindingDB | Database | Binding affinity data for drug targets | 2.9 million measurements; useful for independent testing [16] [17] |
The evidence reviewed demonstrates that PDBbind's structural artifacts and organizational limitations significantly influence both deep learning and traditional docking approaches in protein-ligand pose prediction. The database's quality issues can lead models to memorize erroneous structural patterns, while its standard data splits artificially inflate performance metrics through data leakage. These challenges manifest differently across methodological approaches: deep learning models like AlphaFold 3 achieve remarkable accuracy but show unexpected physical inconsistencies under adversarial testing, while molecular docking methods offer greater robustness to novel targets but generally lower peak performance.
Moving forward, the field requires three key developments: (1) more rigorous benchmarking protocols that control for data leakage and similarity, such as time-split validation and adversarial testing; (2) continued expansion and curation of diverse, high-quality datasets that better represent the true chemical space of drug discovery; and (3) hybrid approaches that leverage the pattern recognition capabilities of deep learning while maintaining the physical plausibility offered by traditional methods. By directly addressing the training data divide, researchers can develop more generalizable and reliable pose prediction methods that accelerate drug discovery rather than simply mastering existing datasets.
The prediction of protein-ligand interactions represents a critical frontier in computational biology and drug discovery. This field is currently defined by two fundamentally distinct approaches: the emerging paradigm of holistic complex prediction exemplified by AlphaFold 3, and the established framework of pose and affinity scoring characteristic of traditional molecular docking methods. AlphaFold 3 represents a transformative shift from specialized prediction tools to a unified deep-learning framework capable of modeling complexes containing proteins, nucleic acids, small molecules, ions, and modified residues simultaneously [3]. In contrast, molecular docking methods primarily focus on predicting ligand binding poses and estimating binding affinities within predefined binding sites, typically treating proteins as relatively rigid structures [1].
This comparison guide objectively evaluates the performance characteristics, methodological foundations, and practical applications of these competing approaches. We examine whether AlphaFold 3's revolutionary architecture translates to consistent practical advantages across diverse drug discovery scenarios, or whether specialized docking methods maintain superiority for specific tasks like affinity prediction and drug-like molecule screening.
Performance Comparison
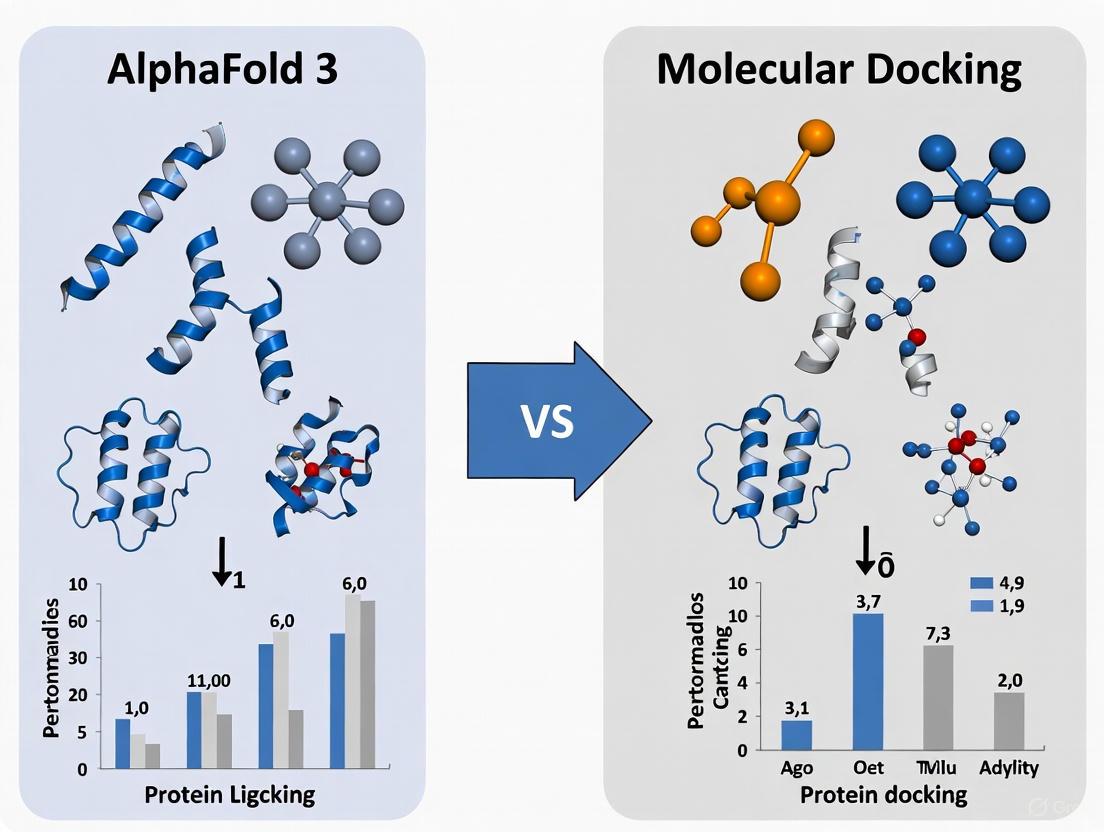
Table 1: Overall performance comparison on the PoseBusters benchmark
| Method | Input Information | PB-Valid Poses (<2Å RMSD) | Key Strengths | Key Limitations |
|---|---|---|---|---|
| AlphaFold 3 (Blind) | Protein sequence + ligand SMILES | 50.3% [4] | Exceptational for blind docking; models full complexes | Lower accuracy on drug-like molecules [4] |
| AlphaFold 3 (Pocket Specified) | Protein sequence + ligand SMILES + pocket residues | 76.6% [4] | High accuracy with minimal structural information | Requires pocket knowledge; commercial use restricted [10] |
| AutoDock Vina (Standard) | Protein structure + ligand | 31.1% [4] | Widely available; fast computation | Lower accuracy on natural ligands [4] |
| Strong Baseline (Vina + Ensemble + Gnina) | Protein structure + ligand + multiple conformations | 69.4% [4] | Superior on drug-like molecules; open access | Requires experimental protein structure [4] |
| DiffDock | Protein structure + ligand | 38% [6] | State-of-the-art prior to AF3 | Lower overall accuracy compared to AF3 [6] |
Specialized Application Performance
Table 2: Performance across specific biological contexts
| Application Domain | Method | Performance Metrics | Context Notes |
|---|---|---|---|
| Antibody-Antigen Docking | AlphaFold 3 (single seed) | 10.2% high-accuracy (DockQ ≥0.8), 34.7% overall success [7] | Improves over AF2-Multimer (2.4% high-accuracy); reaches 60% success with 1000 seeds [7] |
| Nanobody-Antigen Docking | AlphaFold 3 (single seed) | 13.3% high-accuracy, 31.6% overall success [7] | Outperforms Boltz-1 (5%) and Chai-1 (3.33%) on high-accuracy predictions [7] |
| Common Natural Ligands | AlphaFold 3 | Exceptional performance [4] | Molecules highly represented in PDB training data (nucleotides, nucleosides, etc.) [4] |
| Drug-like Molecules (excluding common natural ligands) | Strong Baseline (Vina + Ensemble + Gnina) | 8.5% above AF3 [4] | More representative of typical small-molecule therapeutics [4] |
| Halogenated Compounds (69 PoseBusters ligands) | Strong Baseline | 84.1% PB-valid with RMSD<2Å [4] | Performance on molecules rare in training data |
Methodological Foundations
AlphaFold 3 Architectural Framework
AlphaFold 3 employs a substantially updated diffusion-based architecture that replaces the complex structural module of AlphaFold 2. The system combines a simplified pairformer module with a diffusion network that operates directly on raw atom coordinates, eliminating the need for amino-acid-specific frames and stereochemical violation penalties [3]. The model uses a cross-distillation training approach, enriching training data with structures predicted by AlphaFold-Multimer to reduce hallucination behavior in unstructured regions [3].
The inputs to AlphaFold 3 are notably minimal—requiring only molecular sequences (for proteins, nucleic acids) and SMILES strings (for small molecules)—and the system simultaneously models the complete assembly rather than docking components sequentially [10] [3]. This holistic approach captures the cooperative reshaping that occurs when molecules interact in biological systems.
Traditional Molecular Docking Framework
Traditional docking methods follow a search-and-score paradigm, exploring possible ligand conformations and orientations within a defined binding site, then ranking these poses using scoring functions that estimate binding affinity [1]. These methods exist on a spectrum of flexibility—from rigid-body docking to approaches that allow limited ligand and protein flexibility.
The "strong baseline" approach referenced in Table 1 enhances traditional docking through two key modifications: using ensemble conformations of ligands to ensure adequate sampling of ring geometries and other inflexible regions, and employing machine learning-based rescoring (Gnina) to improve pose selection beyond what traditional scoring functions like Vina provide [4].
Experimental Protocols
PoseBusters Benchmark Methodology
The PoseBusters benchmark established a standardized framework for evaluating protein-ligand complex prediction methods. The test set comprises 428 protein-ligand structures released to the PDB in 2021 or later, ensuring temporal separation from training data for most methods [3]. Evaluation metrics include:
- RMSD <2Å: The classic metric for docking success, measuring heavy-atom root-mean-square deviation after optimal alignment of the binding pocket.
- PB-Valid: A more comprehensive quality check that includes stereochemical validity, absence of severe clashes, and overall physical plausibility [4].
For AlphaFold 3 evaluation, the model was tested in two configurations: truly blind (using only sequence information) and pocket-specified (provided with protein residues constituting the binding site) [4]. Traditional docking methods were evaluated using experimentally determined protein structures.
Adversarial Testing Protocol
Recent research has subjected AlphaFold 3 to adversarial testing to evaluate its understanding of physical principles rather than statistical correlations [6]. The binding site mutagenesis protocol systematically challenges the model:
- Residue Selection: Identify all binding site residues forming contacts with the ligand in the native structure.
- Progressive Mutation:
- Challenge 1: Replace all binding site residues with glycine (removing side-chain interactions)
- Challenge 2: Replace all binding site residues with phenylalanine (steric occlusion)
- Challenge 3: Replace with chemically dissimilar residues (altering shape and chemical properties)
- Evaluation: Measure whether the model adjusts predictions according to physical principles or maintains poses based on training set statistics [6].
Results revealed that co-folding models frequently maintain ligand placement even after removing favorable interactions, indicating potential overfitting to specific system geometries present in training data [6].
Critical Analysis
Physical Realism and Generalization
While AlphaFold 3 demonstrates exceptional accuracy on standard benchmarks, adversarial testing reveals significant limitations in physical understanding. When binding site residues are mutated to glycine, removing key interactions, AlphaFold 3 often continues to predict similar binding poses as if those interactions were still present [6]. In more extreme cases where residues are mutated to phenylalanine, the model sometimes predicts structures with unphysical atomic clashes, indicating difficulty resolving severe steric conflicts within the diffusion process [6].
This suggests that AlphaFold 3's performance derives partly from pattern recognition of complexes in its training set rather than true physical reasoning about molecular interactions. The model appears to learn which ligands tend to bind to particular protein pockets rather than fundamentally understanding how chemical forces dictate binding geometry [6].
Data Dependence and Transferability
Performance analysis reveals substantial variation across molecule types. AlphaFold 3 excels with common natural ligands like nucleotides and cofactors that are well-represented in the Protein Data Bank [4]. However, its advantage diminishes for synthetic drug-like molecules, particularly those containing halogens or other uncommon functional groups [4].
This pattern suggests that data representation in training significantly influences model performance. The "strong baseline" docking approach outperforms AF3 on molecules excluding common natural ligands (69.4% vs 50.3% for blind AF3) [4], indicating that traditional methods may currently be more reliable for typical drug discovery applications involving novel chemical matter.
Practical Considerations for Drug Discovery
For researchers selecting between these approaches, several practical considerations emerge:
- Structural Information Availability: AlphaFold 3 provides remarkable capabilities when experimental protein structures are unavailable, while traditional docking requires high-quality protein structures.
- Computational Resources: AlphaFold 3 demands significant computational resources, especially for large complexes or multiple sampling seeds, whereas traditional docking is relatively lightweight.
- Commercial Applications: AlphaFold 3's current license restricts commercial use, while traditional docking tools are generally open-source or commercially available.
- Dynamic Information: Neither approach adequately captures protein flexibility and dynamics, though traditional docking can be combined with molecular dynamics simulations.
Research Reagent Solutions
Table 3: Essential computational tools for protein-ligand interaction studies
| Tool/Resource | Type | Primary Function | Access Method |
|---|---|---|---|
| AlphaFold Server | Web Server | Holistic complex prediction with minimal input | Free academic access via web interface [10] |
| AutoDock Vina | Software Suite | Traditional molecular docking with empirical scoring | Open-source download [4] |
| Gnina | Software Tool | Machine learning-based pose rescoring | Open-source framework [4] |
| RDKit | Cheminformatics Library | Ligand conformation generation and manipulation | Open-source Python library [4] |
| PoseBusters | Validation Suite | Standardized benchmark for docking methods | Python package [4] |
| PDBBind | Database | Curated protein-ligand complexes for training/testing | Academic license [1] |
The comparison between AlphaFold 3's holistic complex prediction and traditional pose and affinity scoring reveals a nuanced landscape where each approach excels in different scenarios. AlphaFold 3 represents a revolutionary capability for blind prediction of biomolecular complexes, particularly when structural information is limited or for natural biomolecules. However, traditional docking methods, especially when enhanced with machine learning rescoring and conformational ensembles, maintain competitive performance for drug-like molecules and benefit from greater accessibility, speed, and commercial usability.
The optimal approach for research and drug discovery likely involves strategic combination of these technologies—using AlphaFold 3 for initial target assessment and binding site identification, then employing refined docking methods for detailed pose prediction and optimization of novel chemical entities. As both methodologies continue to evolve, the integration of physical principles with data-driven pattern recognition will likely bridge the current gaps, enabling more robust and predictive modeling of protein-ligand interactions across the chemical and biological spectrum.
Performance and Practical Application in Biomolecular Modeling
The accurate prediction of how a small molecule (ligand) binds to its target protein is a cornerstone of modern drug discovery. For years, classical docking tools like AutoDock Vina have been the standard for this task. The recent release of AlphaFold 3 (AF3), a deep learning model capable of predicting protein-ligand complexes from sequence alone, promises a paradigm shift [3]. This guide provides an objective comparison of the docking accuracy between AF3 and traditional molecular docking methods, focusing on the critical metrics of ligand Root-Mean-Square Deviation (RMSD) and success rates on standard benchmarking datasets. The analysis is framed within the broader thesis of evaluating the role of AI-driven versus physics-based methods in structural bioinformatics.
Quantitative Performance Comparison
The performance of a docking tool is primarily measured by its ability to produce a ligand pose that is close to the experimentally determined structure. A common threshold for a "successful" prediction is a ligand RMSD of less than 2.0 Å when the predicted pose is aligned to the protein's binding pocket.
The table below summarizes the performance of AF3 and various docking methods on the PoseBusters benchmark, a curated set of protein-ligand structures released after AF3's training data cutoff, ensuring an unbiased evaluation [21] [4].
Table 1: Success Rate (% of complexes with pocket-aligned ligand RMSD < 2.0 Å) on the PoseBusters Benchmark
| Method | Input Type | Reported Success Rate | Notes |
|---|---|---|---|
| AlphaFold 3 (Blind) | Protein Sequence + Ligand SMILES | ~48% | No protein structure input [3] [4] |
| AlphaFold 3 (Pocket Specified) | Protein Sequence + Ligand SMILES + Pocket Residues | ~62% | Protein residues near the ligand are specified [4] |
| AutoDock Vina (Baseline) | Protein Structure + Ligand Structure | ~33% | As reported in PoseBusters and AF3 papers [22] [3] |
| Strong Baseline (Vina + Ensembles + Gnina) | Protein Structure + Ligand Structure | ~52% | Uses an ensemble of ligand conformations & neural network rescoring [4] |
Performance can vary significantly with the type of ligand being docked. AF3 demonstrates particular strength on "common natural ligands" (e.g., nucleotides), which are well-represented in its training data. In contrast, traditional docking shows more consistent performance across diverse, drug-like molecules [4].
Table 2: Performance on Different Ligand Types within the PoseBusters Benchmark
| Method | Common Natural Ligands (n=50) | Other Ligands (More Drug-like) |
|---|---|---|
| AlphaFold 3 (Blind) | Higher Performance | Lower Performance |
| Strong Docking Baseline | Lower Performance | ~8.5% higher than blind AF3 |
Beyond general small molecules, benchmarking on specific pollutant compounds like Per- and polyfluoroalkyl substances (PFAS) reveals another nuance. AF3's performance was notably higher on data it was trained on ("Before Set": ~74.5% success) compared to unseen data ("After Set": ~55.8% success), indicating potential overfitting. A hybrid approach, using AF3 to identify the binding pocket and Vina for the final pose prediction, proved to be a successful strategy [22].
Experimental Protocols and Metrics
Standard Benchmarking Datasets
The reliability of any performance claim hinges on the use of rigorous, non-overlapping datasets.
- PoseBusters Benchmark: This is a key modern benchmark comprising 428 protein-ligand complexes from the PDB released in 2021 or later. It is designed to test methods on structures not used in their training, providing a fair assessment of generalizability [3] [21].
- Curated PDB Sets: Studies often create their own benchmarks from the Protein Data Bank (PDB). A critical practice is splitting data into "Before" and "After" sets based on a model's training cutoff date (e.g., September 30, 2021, for AF3) to evaluate performance on seen versus unseen data [22].
- Directory of Useful Decoys (DUD): While primarily used for evaluating virtual screening enrichment (distinguishing binders from non-binders), the principles of DUD—using physically similar but chemically distinct decoys—inform the creation of stringent docking benchmarks [23].
Key Performance Metrics
- Ligand RMSD: The most direct metric for pose accuracy. It measures the average distance between the atoms of the predicted ligand pose and the experimentally determined (native) pose after aligning the protein's binding pocket atoms. A lower RMSD indicates a more accurate prediction.
- Success Rate: The percentage of complexes in a benchmark for which the ligand RMSD is below a chosen threshold (typically 2.0 Å).
- Protein-Ligand Interaction Fidelity (PLIF): Beyond RMSD, recovering the specific hydrogen bonds, hydrophobic contacts, and other interactions found in the native structure is crucial. A pose with good RMSD may still have incorrect interactions, which can mislead drug design. Studies show that classical docking methods, whose scoring functions explicitly seek these interactions, often achieve better PLIF recovery than some ML methods that only use RMSD-derived loss functions [21].
Method Workflows
The workflow for benchmarking varies significantly between AF3 and classical docking tools.
The Scientist's Toolkit: Essential Research Reagents
To conduct a rigorous docking benchmark, researchers require both software tools and carefully curated data.
Table 3: Key Reagents for Docking Benchmarking Studies
| Reagent / Resource | Type | Function in Benchmarking | Example |
|---|---|---|---|
| Benchmarking Datasets | Data | Provides standardized, non-overlapping complexes for fair evaluation of method performance and generalizability. | PoseBusters Benchmark [21], PDB "After Sets" [22] |
| Structure Preparation Tools | Software | Prepares protein and ligand structures for docking by adding hydrogens, assigning charges, and minimizing conflicts. | PDBFixer [22], OpenBabel [22], Spruce (OpenEye) [21] |
| Classical Docking Suites | Software | Provides physics-inspired or knowledge-based algorithms for conformational sampling and pose scoring. | AutoDock Vina [22], Gnina [4], GOLD [21] |
| AI-Based Prediction Tools | Software | Predicts complex structures end-to-end from sequence and SMILES string, often with high speed. | AlphaFold 3 Server [3], DiffDock-L [21] |
| Interaction Analysis Packages | Software | Analyzes and compares predicted poses against ground truth by calculating interaction fingerprints. | ProLIF [21] |
| Analysis Metrics | Scripts/Metrics | Quantifies the accuracy of predicted poses through structural alignment and interaction recovery. | RMSD, Success Rate, Protein-Ligand Interaction Fidelity (PLIF) [21] |
The benchmarking data leads to several key conclusions for researchers:
- AlphaFold 3's Niche: AF3 is a revolutionary tool for blind docking, where a protein structure is unavailable, achieving remarkable accuracy using only sequence information. It performs exceptionally well on ligand types common in its training data.
- Classical Docking's Resilience: When a high-quality experimental protein structure is available, strengthened classical docking pipelines that use conformational ensembles and machine learning-based rescoring (e.g., with Gnina) can match or even exceed the performance of the blind version of AF3, especially on drug-like molecules.
- The Power of Hybrids: A promising strategy is a hybrid approach, using AF3's strengths in pocket identification and then refining the pose with physics-based tools like Vina, which has shown improved results for challenging molecules like PFAS [22].
- Look Beyond RMSD: For critical drug discovery applications, evaluating protein-ligand interaction fingerprints (PLIF) is essential, as a good RMSD does not guarantee the recovery of key biochemical interactions [21].
In summary, AF3 has not rendered traditional docking obsolete but has instead expanded the toolkit. The choice between them is context-dependent. For the foreseeable future, integrating the predictive power of deep learning with the physicochemical rigor of classical methods will likely provide the most robust and reliable strategy for protein-ligand pose prediction.
The accurate computational prediction of how biomolecules interact is a cornerstone of modern drug discovery and basic biological research. For years, molecular docking—a physics-inspired method that leverages known protein structures to predict where and how small molecules bind—has been the dominant technique. The recent emergence of deep learning systems like AlphaFold 3 (AF3) represents a paradigm shift, offering a unified approach to predicting the joint 3D structures of diverse biomolecular complexes directly from their sequence information. This guide provides an objective comparison of AlphaFold 3 and traditional molecular docking for predicting the structures of proteins, antibodies, and nanobodies with their molecular partners, synthesizing current performance data and detailing key experimental methodologies.
AF3 employs a substantially updated architecture compared to its predecessors, capable of predicting the joint structure of complexes including proteins, nucleic acids, small molecules, ions, and modified residues. Its core innovation lies in a diffusion-based approach that starts with a cloud of atoms and iteratively refines the most probable molecular structure, operating directly on raw atom coordinates without the need for complex rotational adjustments [3] [24]. This allows AF3 to handle arbitrary chemical components while maintaining chemical plausibility. In contrast, traditional docking tools like Vina rely on physics-based scoring functions and require an experimentally determined protein structure as a starting point, which can be a significant limitation in early-stage research [4].
Performance Comparison: AlphaFold 3 vs. Molecular Docking
The most cited benchmark for protein-ligand docking is the PoseBusters set, comprising 428 protein-ligand structures released to the PDB in 2021 or later. The results demonstrate AF3's strong performance, particularly given that it operates without structural inputs.
Table 1: Protein-Ligand Docking Accuracy on the PoseBusters Benchmark
| Method | Input Requirements | PB-Valid & RMSD <2Å (%) | Notes |
|---|---|---|---|
| AlphaFold 3 (Blind) | Protein sequence, Ligand SMILES | 26.3% | No structural information used [3] |
| AlphaFold 3 (Pocket Specified) | Protein sequence, Ligand SMILES, Protein residues near ligand | 33.6% | Still uses sequence, not 3D structure [4] |
| Vina (Baseline) | Experimental protein structure, Ligand | 11.1% | Original baseline from PoseBusters paper [4] |
| Strong Baseline (Vina + Gnina) | Experimental protein structure, Ligand conformational ensemble | 30.3% | Combines ensemble docking & neural network rescoring [4] |
A critical analysis reveals that while the AF3 paper showed it "greatly outperforms classical docking tools like Vina," the Vina baseline does not represent the state-of-the-art in traditional docking. When strengthened with standard improvements—using an ensemble of ligand starting conformations and rescoring poses with the neural network-based Gnina—the performance of traditional docking nearly matches that of the pocket-specified version of AF3 [4]. This strong baseline uses a earlier training data cutoff than AF3, ensuring a fair comparison.
Performance varies significantly by ligand type. AF3 demonstrates exceptional performance on "common natural ligands" (e.g., nucleosides, nucleotides), which are highly represented in its training data due to their frequent occurrence in the PDB. However, the strengthened baseline outperforms AF3 on the remaining molecules, which may be more representative of typical drug-like compounds [4].
Antibody and Nanobody Complex Prediction
Antibody and nanobody docking presents a unique challenge due to the flexibility of their complementary-determining regions (CDRs), particularly the highly diverse CDR H3 loop. Accurate prediction here is critical for therapeutic development.
Table 2: Antibody and Nanobody Docking Success Rates (DockQ ≥0.23)
| Method | Antibody-Antigen Success Rate | Nanobody-Antigen Success Rate | Sampling Conditions |
|---|---|---|---|
| AlphaFold 3 | 34.7% | 31.6% | Single seed [7] |
| AlphaFold 3 (with 1000 seeds) | ~60% | Not reported | Extensive sampling [7] |
| AlphaFold 2.3-Multimer | 23.4% | Not reported | Standard [7] |
| Boltz-1 (AF3-like) | 20.4% | 23.3% | Single seed [7] |
| Chai-1 (AF3-like) | 20.4% | 15.0% | Single seed [7] |
| AlphaRED (Hybrid) | 43% | Not reported | Combines AF2 with replica exchange docking [25] |
AF3 shows a clear improvement over AF2-Multimer, but its success rate with a single seed remains limited at 34.7%. However, its performance can nearly double with extensive sampling (1,000 seeds), highlighting the stochastic nature of the diffusion model [7]. This comes at a significant computational cost. The hybrid method AlphaRED, which combines AF2 structural templates with physics-based replica exchange docking, achieves a higher success rate on antibody-antigen targets, demonstrating the value of integrating deep learning with physics-based sampling [25].
For nanobodies, the overall success rate of both AF3 and AF2-Multimer remains below 50%, though AF3 shows a modest overall improvement. Accuracy is heavily influenced by the characteristics of the CDR3 loop, particularly its 3D spatial conformation and length [26].
Linear Epitope Prediction with AlphaFold-based Pipelines
For predicting linear antibody epitopes (short, contiguous peptide sequences bound by antibodies), specialized pipelines built upon AlphaFold2 have been developed. The PAbFold pipeline uses the localColabFold implementation of AF2 to predict the structure of a single-chain variable fragment (scFv) in complex with overlapping peptides derived from an antigen [27] [28]. This method has been experimentally validated to accurately flag known epitope sequences for well-characterized antibodies and for a novel anti-SARS-CoV-2 antibody, with predictions verified via peptide competition ELISA [28]. The computational expense scales with the square of the concatenated sequence length, making the use of minimized scFvs and short peptides efficient (approximately 1.5 minutes per scFv-peptide complex on an NVIDIA A5000 GPU) [27].
Experimental Protocols and Methodologies
Standard AlphaFold 3 Protocol for Complex Prediction
The standard workflow for using AF3 to model a biomolecular complex involves several key steps. The required inputs are the sequences of all polymeric components (e.g., protein, DNA, RNA) and the SMILES string for any small molecule ligands. The process is managed through the AlphaFold Server, which is designed to be accessible to scientists.
The model's architecture begins by processing inputs through a simplified Multiple Sequence Alignment (MSA) module, which is substantially de-emphasized compared to AlphaFold 2. The "Pairformer" module then evolves a pairwise representation of the entire complex. Finally, the diffusion module, which replaces the structure module of AF2, generates atomic coordinates through an iterative denoising process [3]. A critical technical point is that AF3 uses a cross-distillation method during training, where it is trained on structures predicted by AlphaFold-Multimer. This teaches the model to represent unstructured regions as extended loops rather than compact hallucinations, greatly reducing a common failure mode of generative models [3].
Strong Baseline Docking Protocol
The strengthened traditional docking baseline, which performs comparably to AF3, can be implemented in approximately 100 lines of code and uses open-source tools [4]. The following diagram illustrates this integrated workflow, which combines the strengths of deep-learning initial sampling with physics-based refinement and selection.
Key Steps:
- Generate Conformational Ensemble: Using a cheminformatics toolkit like RDKit, generate multiple reasonable 3D starting conformations for the ligand from its SMILES string. This ensures the docking algorithm can sample correct poses even for rigid ring systems [4].
- Molecular Docking: Run the Vina docking software from each starting ligand conformation against the experimental protein structure. The exhaustiveness parameter can be reduced for each run since the ensemble provides broader sampling.
- Rescore and Select Top Poses: Pool all docked poses from the various runs and rescore them using Gnina, a convolutional neural network trained to distinguish near-native docking poses. Select the top-ranked pose based on the Gnina score, which is more accurate than Vina's native scoring function [4].
Protocol for Antibody-Antigen Docking with AlphaRED
The AlphaRED protocol is a hybrid approach that addresses the limitations of AF models for docking antibodies and other flexible complexes [25].
Workflow:
- Generate Structural Templates: Use AlphaFold-Multimer to generate a diverse set of structural models for the antibody-antigen complex.
- Estimate Flexibility and Interface Quality: Repurpose AF's confidence metrics (pLDDT and predicted aligned error) to estimate protein flexibility and identify which template models have the most reliable interfaces.
- Physics-Based Refinement: Use the best AF-generated models as starting points for the RosettaDock replica exchange docking protocol. This physics-based method extensively samples side-chain conformations, backbone flexibility, and rigid-body degrees of freedom to refine the complex.
- Select Final Models: Rank the resulting models using a combination of Rosetta's energy function and the original AF confidence metrics to produce the final, high-accuracy predictions [25].
Table 3: Key Software and Data Resources for Biomolecular Modeling
| Resource Name | Type | Function and Application |
|---|---|---|
| AlphaFold Server | Web Server | Free, accessible interface for running AlphaFold 3 predictions on biomolecular complexes [24]. |
| PoseBusters Benchmark | Dataset & Software | A benchmark set of 428 protein-ligand complexes and a Python package to validate docking poses, ensuring they are <2Å from experimental structures and free of stereochemical violations [4]. |
| Gnina | Software | A molecular docking software that uses a convolutional neural network to score and select the most accurate docking poses from a pool of candidates [4]. |
| RDKit | Software | An open-source cheminformatics toolkit used to generate and manipulate small molecule structures, including the creation of conformational ensembles [4]. |
| SAbDab | Database | The Structural Antibody Database, a repository of all publicly available antibody structures, used for curating benchmark sets [7]. |
| PAbFold | Software Pipeline | A computational pipeline based on AlphaFold2 and localColabFold for predicting linear antibody epitopes by modeling scFv-peptide complexes [27] [28]. |
| AlphaRED | Software Pipeline | A hybrid pipeline integrating AlphaFold with Rosetta-based replica exchange docking for reliable protein-protein and antibody-antigen docking [25]. |
The comparison between AlphaFold 3 and molecular docking reveals a nuanced landscape. AF3 is a breakthrough for blind docking, achieving high accuracy using only sequence information where traditional methods require a known protein structure. This makes it invaluable for targets with no experimentally determined structure. However, when a high-quality experimental structure of the target protein is available, strengthened traditional docking baselines can achieve comparable, and in some cases superior, accuracy, especially for drug-like molecules [4].
For antibody and nanobody docking, AF3 represents a step forward, but challenges remain. Its single-seed success rate is modest, and achieving high accuracy often requires computationally expensive massive sampling. Hybrid approaches like AlphaRED, which combine deep learning's sampling power with physics-based refinement, currently set the state-of-the-art for these difficult targets [25].
The choice between these tools is therefore context-dependent. For rapid, initial assessment of a novel target, AF3 is unparalleled. For optimizing drug candidates against a well-characterized target with an available structure, strengthened traditional docking or hybrid methods may provide superior results. The future of biomolecular modeling lies not in a single tool dominating, but in the intelligent integration of these complementary approaches to accelerate scientific discovery and therapeutic development.
The accurate prediction of biomolecular structures is a cornerstone of modern drug discovery and basic biological research. For years, molecular docking has been the predominant computational method for predicting how small molecules interact with their protein targets. However, the recent advent of deep learning-based cofolding tools, like AlphaFold 3 (AF3), represents a paradigm shift. This guide provides an objective comparison of AlphaFold 3 and traditional molecular docking, focusing on their performance in predicting the poses of ligands bound to challenging target classes: RNA, membrane proteins, and proteins with flexible loops. We summarize quantitative data from recent benchmarks and detail key experimental protocols to help researchers select the appropriate tool for their pose prediction challenges.
The table below summarizes the core strengths and weaknesses of AlphaFold 3 and molecular docking across key biomolecular categories, synthesizing findings from recent evaluations [3] [10] [29].
Table 1: Comparative Performance of AlphaFold 3 vs. Molecular Docking
| Target Category | AlphaFold 3 Performance | Molecular Docking Performance | Key Supporting Evidence |
|---|---|---|---|
| Overall Protein-Ligand Pose Prediction | High performance, often doubling the accuracy of traditional docking; excels in "blind" scenarios using only sequence/SMILES [3] [10]. | Variable and often lower, especially without a pre-defined holo structure; performance can be improved with fragment-derived priors or in "easy" splits [30] [29] [31]. | On the PoseBusters benchmark, AF3 significantly outperformed docking tools like Vina, with a much higher percentage of predictions within 2 Å RMSD [3]. |
| RNA Structures | Mixed to poor; identified as a weakness due to RNA's conformational flexibility [10]. | Not typically used for full RNA-ligand co-structure prediction. | AF3 struggles with RNA's context-dependent folding, and predictions in this area require extra skepticism [10]. |
| Membrane Proteins | Challenging; the model does not explicitly account for lipid bilayers, leading to potential artifacts in transmembrane regions [10]. | Performance is highly dependent on the quality and state (e.g., apo vs. holo) of the input protein structure [29]. | Critical drug targets like GPCRs modeled by AF3 need careful interpretation due to the lack of a membrane environment [10]. |
| Proteins with Flexible Loops | Can identify disordered regions but cannot predict their dynamic behavior [10]. | Performance can be poor if the loop conformation in the input structure differs significantly from the bound state (e.g., due to "induced fit") [29]. | In high-throughput docking benchmarks, even small side-chain variations in AF models compared to experimental structures consistently reduced performance [29]. |
Detailed Experimental Protocols and Methodologies
The PoseBusters Benchmark (For AF3 and Docking)
The PoseBusters benchmark has become a standard for rigorously evaluating protein-ligand pose prediction methods [3].
- Objective: To assess the ability of a method to generate a ligand pose that matches the experimental structure, measured by the root-mean-square deviation (RMSD) of the ligand heavy atoms after aligning the protein pocket.
- Dataset: Consists of 428 protein-ligand structures released to the PDB in 2021 or later. This time-split is crucial for testing generalizability and avoiding data leakage from the training sets of data-driven models like AF3 [3].
- Metric: The primary metric is the percentage of protein-ligand pairs with a pocket-aligned ligand RMSD of less than 2 Å, which is a common threshold for a "successful" prediction.
- Key Findings:
- AlphaFold 3: Demonstrated substantially higher accuracy than state-of-the-art docking tools, even though AF3 uses only the protein sequence and ligand SMILES string, while docking methods often use the experimental protein structure as input [3].
- Molecular Docking: Its performance is often lower in this blind setting. However, its accuracy can be boosted by incorporating data-driven priors. For example, one workflow using Vina-GPU augmented with fragment-derived priors achieved over 50% success for SARS-CoV-2 and MERS-CoV protease targets [31].
High-Throughput Docking (HTD) on AlphaFold Models
This protocol evaluates the direct utility of predicted protein structures for virtual screening [29].
- Objective: To determine if an AlphaFold-predicted protein structure can reliably replace an experimental one in a docking-based virtual screening campaign to identify new active molecules.
- Dataset: A benchmark set of 22 diverse protein targets, comparing AF models from the AlphaFold Database with their corresponding experimental PDB structures [29].
- Methodology:
- Structure Preparation: Both PDB and AF structures are stripped of water, ions, and co-factors to ensure a fair comparison.
- Docking: Multiple docking programs (e.g., AutoDock 4, ICM, rDock, PLANTS) are used to screen a library of known actives and decoys.
- Evaluation: Performance is measured by the enrichment factor—the ability to rank active molecules early in the list—and the success in recapitulating the pose of the native ligand.
- Key Findings:
- AF models showed consistently worse HTD performance compared to experimental structures, despite having good overall structural accuracy (low backbone RMSD) [29].
- Even small side-chain variations in the binding site, particularly in flexible loops, were sufficient to significantly impact docking accuracy and enrichment [29].
Visualizing the Methodologies
The fundamental difference between AF3 and docking lies in their approach. AF3 is a cofolding method that predicts the entire complex simultaneously, while docking is a sequential process that relies on a pre-existing protein structure.
Diagram 1: Cofolding vs. Sequential Docking Workflows
The table below lists key software tools and databases mentioned in this guide that are essential for conducting rigorous pose prediction research.
Table 2: Key Reagents and Resources for Pose Prediction Research
| Resource Name | Type | Primary Function in Research | Relevance to Comparison |
|---|---|---|---|
| AlphaFold Server | Web Server | Free academic access to AlphaFold 3 for predicting structures of protein-ligand complexes [10]. | Primary tool for generating AF3 predictions for a target of interest. |
| AlphaFold Protein Structure Database | Database | Repository of pre-computed AF and AF3 structures for a vast number of proteins [29]. | Source of "as-is" AF models for docking studies without running the predictor. |
| PDB (Protein Data Bank) | Database | The primary global archive for experimentally determined 3D structures of biological macromolecules [32]. | Source of ground-truth structures for benchmarking and validation. |
| PoseBusters Benchmark | Benchmark Suite | A set of tests to validate the physical realism and geometric correctness of predicted molecular complexes [3]. | Standardized benchmark for evaluating pose prediction method performance. |
| RDKit | Software Library | An open-source toolkit for cheminformatics, used for ligand handling, MCS detection, and conformer generation [30]. | Core utility in many computational chemistry workflows, including the TEMPL baseline method [30]. |
| Vina-GPU | Software Tool | An open-source docking program accelerated for GPUs, used with data-driven priors [31]. | Representative of traditional docking methods used in modern, augmented workflows. |
The introduction of AlphaFold 3 (AF3) represents a paradigm shift in computational structural biology, moving beyond traditional molecular docking through its unified deep learning framework for modeling biomolecular complexes. This comparison guide objectively evaluates AF3 against established docking methods and emerging alternatives, examining their integration into real-world drug discovery and antibody design pipelines through published performance metrics and experimental protocols.
Performance Benchmarking: Quantitative Comparison
Table 1: Protein-Ligand Docking Performance Comparison
| Method | Type | Accuracy (Ligand RMSD < 2Å) | Benchmark | Sampling Conditions |
|---|---|---|---|---|
| AlphaFold 3 | Co-folding | 81% (blind), 93% (with site) | PoseBusterV2 | Default server settings [6] |
| DiffDock | Deep learning docking | 38% | PoseBusterV2 | Not specified [6] |
| AutoDock Vina | Traditional docking | ~60% | PoseBusterV2 | With known binding site [6] |
| RoseTTAFold All-Atom | Co-folding | Lower than AF3 (exact % not specified) | PoseBusterV2 | Default settings [6] |
| Pearl (Genesis) | Co-folding | ~15% improvement over AF3 | Runs N' Poses | Not specified [33] |
Antibody and Nanobody Docking Performance
Table 2: Antibody-Antigen Complex Prediction Accuracy
| Method | High-Accuracy Success (Antibodies) | High-Accuracy Success (Nanobodies) | Sampling Conditions | Benchmark |
|---|---|---|---|---|
| AlphaFold 3 | 10.2% | 13.3% | Single seed [7] | Curated Ab/Ag benchmark |
| AlphaFold 3 (reported by DeepMind) | 60% | Not specified | 1,000 seeds [7] | Internal benchmark |
| AF2.3-Multimer | 2.4% | Not specified | Standard sampling [7] | Curated Ab/Ag benchmark |
| Boltz-1 | 4.1% | 5.0% | Single seed, 3 recycles [7] | Curated Ab/Ag benchmark |
| Chai-1 | 0% | 3.3% | Single seed, 3 recycles [7] | Curated Ab/Ag benchmark |
| AlphaRED (AF2.3-M + Rosetta) | 43% | Not specified | Standard sampling [7] | Curated Ab/Ag benchmark |
| Traditional Rosetta docking | 20% | Not specified | Standard sampling [7] | CAPRI standards |
Methodologies and Experimental Protocols
AlphaFold 3 Architecture and Workflow
AF3 employs a substantially updated diffusion-based architecture that replaces AlphaFold 2's structure module. The system uses a pairformer block that de-emphasizes multiple sequence alignment (MSA) processing in favor of direct atomic coordinate prediction through a diffusion process [3]. During inference, AF3 starts with random noise and iteratively refines atomic positions through a denoising process that learns to generate biologically plausible structures [3] [10].
The model is trained on nearly all structural data in the Protein Data Bank, incorporating proteins, nucleic acids, small molecules, ions, and modified residues within a single unified framework. A critical technical innovation is the cross-distillation method that enriches training data with structures predicted by AlphaFold-Multimer to reduce hallucination in unstructured regions [3].
Traditional Molecular Docking Protocol
Traditional docking methods like AutoDock Vina primarily follow a search-and-score framework [1]. The standard workflow involves:
- Preparation: Isolating the protein receptor and ligand structures, adding hydrogen atoms, and assigning partial charges.
- Search Algorithm: Exploring the conformational space of the ligand within the binding site using algorithms like Monte Carlo, genetic algorithms, or molecular dynamics.
- Scoring Function: Evaluating each generated pose using physics-inspired or empirical scoring functions to estimate binding affinity.
- Post-processing: Clustering similar poses and selecting top candidates based on scoring function values [1].
These methods typically treat proteins as rigid bodies or allow limited side-chain flexibility, balancing computational efficiency against accuracy [1].
RFdiffusion for Antibody Design
The RFdiffusion protocol for de novo antibody design involves fine-tuning the network specifically on antibody complex structures [34]. The methodology includes:
- Conditioning: Providing the antibody framework structure and sequence as input while allowing the network to design complementarity-determining regions (CDRs).
- Epitope Targeting: Using one-hot encoded "hotspot" features to direct antibodies toward specific epitopes.
- Rigid-body Placement: Designing both CDR loop conformations and the overall orientation of the antibody relative to the target.
- Sequence Design: Using ProteinMPNN to design CDR loop sequences after structural generation [34].
This approach has demonstrated atomic-level accuracy in designing antibody variable heavy chains (VHHs) and single-chain variable fragments (scFvs) targeting disease-relevant epitopes, with cryo-EM validation confirming design accuracy [34].
Workflow Integration Diagrams
Table 3: Key Computational Tools and Experimental Methods
| Tool/Resource | Type | Primary Function | Application Context |
|---|---|---|---|
| AlphaFold Server | Web service | Biomolecular complex prediction | Academic research, non-commercial use [10] |
| RFdiffusion | Software | De novo protein and antibody design | Epitope-specific antibody generation [34] |
| ProteinMPNN | Software | Protein sequence design | Designing sequences for RFdiffusion structures [34] |
| PoseBusterV2 | Benchmark dataset | Method validation for protein-ligand docking | Performance evaluation [6] |
| AutoDock Vina | Software | Traditional molecular docking | Baseline comparisons, hybrid workflows [6] [1] |
| SAbDab | Database | Structural antibody data | Benchmarking antibody-specific methods [7] |
| Yeast Surface Display | Experimental system | High-throughput antibody screening | Validation of computational designs [34] |
| Surface Plasmon Resonance | Experimental system | Binding affinity measurement | Kinetic characterization of designs [34] |
Limitations and Practical Considerations
Physical Realism and Robustness
Recent adversarial testing reveals significant limitations in co-folding models' understanding of physical principles. When binding site residues in Cyclin-dependent kinase 2 (CDK2) were mutated to glycine or phenylalanine, AF3 and similar models continued to place ATP in the original binding site despite the loss of favorable interactions and introduction of steric clashes [6]. This indicates potential overfitting to training data rather than genuine learning of physical interactions.
Context Dependencies and Failures
Performance varies substantially across biomolecular types. While AF3 demonstrates strong protein-ligand prediction capabilities, RNA structure prediction remains challenging due to conformational flexibility [10]. For antibody docking, approximately 65% of predictions fail to achieve correct docking even with single-seed sampling, indicating substantial room for improvement [7].
Glycan modeling presents particular challenges, as correct stereochemistry preservation is highly context-dependent and requires specialized input formats like Bonded AtomPairs (BAP) syntax for accurate predictions [35].
Accessibility and Implementation
AF3's initial release limited access to a web server with non-commercial restrictions, though academic code and weights were subsequently released [10]. This contrasts with more open traditional docking tools and creates barriers for commercial drug discovery applications. Integration into automated pipelines may be challenged by server-based access models compared to locally installed traditional tools.
Emerging Alternatives and Future Directions
New models like Pearl (Genesis Molecular AI) claim ~15% improvement over AF3 on the Runs N' Poses benchmark, utilizing large-scale physics-generated synthetic data and SO(3)-equivariant diffusion architectures [33]. These approaches aim to address data scarcity through synthetic training complexes while maintaining physical plausibility.
The integration of co-folding predictions with physics-based refinement represents a promising hybrid approach. Many organizations now use AF3 predictions as starting points for molecular dynamics simulations and binding affinity calculations [10] [33], leveraging the strengths of both deep learning and physics-based methods.
For antibody design, the combination of RFdiffusion structural generation with experimental screening platforms like yeast display enables complete in silico to in vitro workflows [34], potentially accelerating therapeutic antibody development against emerging targets like SARS-CoV-2 variants [36].
Navigating Limitations and Optimizing Prediction Workflows
A critical evaluation of biomolecular structure prediction tools reveals a significant trade-off: while deep learning models like AlphaFold 3 achieve remarkable speed and overall accuracy, their architectural choices can sometimes come at the cost of strict physical realism. Concurrently, modern physics-based docking methods, when properly configured, remain highly competitive, especially in handling drug-like molecules and avoiding steric violations. This guide objectively compares the performance of AlphaFold 3 against other co-folding models and traditional docking approaches on the critical metrics of steric clashes and bond geometry.
Architectural Foundations and Their Impact on Physical Realism
The core architecture of a prediction model fundamentally dictates its approach to maintaining physical realism.
AlphaFold 3 replaces the traditional structure module of its predecessor with a diffusion-based architecture that directly predicts raw atom coordinates [3]. A key innovation is the removal of explicit stereochemical loss functions and complex rotational frame representations, relying instead on the multiscale nature of the diffusion process to learn local stereochemistry [3]. This approach simplifies the handling of diverse chemical components but places the entire burden of learning correct bond geometry on the training data and diffusion process.
In contrast, traditional docking tools like AutoDock Vina are built on physics-inspired scoring functions that explicitly evaluate terms for steric clashes, hydrogen bonding, and hydrophobic interactions [4]. They operate on input structures that typically already have correct bond lengths and angles, thus avoiding the problem of poor bond geometry altogether.
Experimental Evidence from Adversarial Challenges
Rigorous testing through biologically plausible adversarial examples provides critical insights into the physical understanding of co-folding models.
Binding Site Mutagenesis Challenge
A seminal study investigated model robustness by mutating all binding site residues of Cyclin-dependent kinase 2 (CDK2) in complex with ATP to glycine and subsequently to phenylalanine [6]. The results probe the model's reliance on statistical correlations versus physical principles.
- Workflow of the Binding Site Mutagenesis Experiment: The diagram below illustrates the experimental protocol for challenging co-folding models.
- Key Findings:
- Glycine Mutant: All four tested co-folding models (AF3, RFAA, Boltz-1, Chai-1) continued to place the ATP molecule in the original binding site, despite the removal of all major side-chain interactions that originally stabilized the pose [6].
- Phenylalanine Mutant: When the binding site was packed with bulky phenylalanine residues, the models showed some capacity to adapt. However, the predictions remained heavily biased towards the original binding site, with several models producing outputs containing "unphysical overlapping atoms and large steric clashes" [6].
This indicates that while these models learn strong statistical preferences for specific binding pockets, their internal representation does not fully enforce fundamental physical constraints against atomic overlaps, especially when presented with highly unnatural sequences.
Performance Benchmarking on Standardized Tasks
Standardized benchmarks offer a quantitative comparison of model performance on realistic prediction tasks.
Antibody-Antigen Docking Accuracy
The accuracy of CDR H3 loop prediction is a major determinant of success in antibody-antigen docking. Benchmarking on a curated, redundancy-filtered dataset reveals the performance of various models with a single seed [7].
Table 1: Docking Success Rates on Antibody-Antigen Complexes (Single Seed)
| Model | High-Accuracy Success (DockQ ≥0.80) | Overall Success (DockQ >0.23) | Key Observation |
|---|---|---|---|
| AlphaFold 3 (AF3) | 10.2% | 34.7% | Sets a new benchmark for a single, unrefined prediction [7]. |
| AF2.3-Multimer | 2.4% | 23.4% | Serves as a reference for the previous generation [7]. |
| Boltz-1 | 4.1% | 20.4% | An AF3-like model; performance is sensitive to recycling and MSA depth [7]. |
| Chai-1 | 0% | 20.4% | Another AF3-like model; struggled with high-accuracy predictions in this test [7]. |
| AlphaRED | ~43% (with refinement) | N/A | A hybrid method using AF2.3-Multimer + replica exchange docking, showing the value of combining AI with physics-based sampling [25]. |
The data shows that while AF3 represents a significant step forward, its failure rate for antibody docking with a single seed remains high at 65%, underscoring the need for further improvement and/or extensive sampling [7].
Protein-Ligand Docking and Pose Validation
The PoseBusters benchmark, which validates poses for both RMSD accuracy and physical chemical sanity (e.g., steric clashes, bond lengths), is a standard for protein-ligand docking.
- AlphaFold 3 Performance: In its blind docking mode (no protein structure provided), AF3 achieved a ~15% absolute improvement in generating PB-valid poses compared to a standard Vina baseline [4]. When provided with pocket information, this improvement increased to ~26% [4].
- Stronger Baselines: A study demonstrated that a stronger baseline docking pipeline, incorporating ligand conformational ensembles and CNN-based rescoring (Gnina), could outperform the blind version of AF3 by 4.2% on the full PoseBusters set. This baseline came within 7.1% of the pocket-informed AF3 results [4]. Notably, this baseline excelled on molecules excluding common natural ligands, a set potentially more representative of drug-like compounds [4].
Table 2: Comparison of Pose Prediction Methods and Characteristics
| Method / Characteristic | AlphaFold 3 (Blind) | AlphaFold 3 (Pocket-Informed) | Strong Baseline (Vina + Ensembles + Gnina) |
|---|---|---|---|
| Input Requirements | Protein sequence, Ligand SMILES | Protein sequence, Ligand SMILES, Pocket residues | Protein 3D structure, Ligand SMILES |
| PoseBusters Benchmark (PB-valid & <2Å) | ~15% over Vina [4] | ~26% over Vina [4] | ~19% over Vina [4] |
| Performance on Drug-like Molecules | Unclear from public data | Unclear from public data | 8.5% higher than blind AF3 on non-natural ligands [4] |
| Handling of Bond Geometry | Learned implicitly via diffusion; generally good but not explicitly constrained [3] | Learned implicitly via diffusion; generally good but not explicitly constrained [3] | Input ligand conformers have correct geometry; docking does not alter bonds. |
| Typical Steric Clashes | Can occur, as evidenced in adversarial tests [6] | Can occur, as evidenced in adversarial tests [6] | Scoring function includes steric clash term. |
The Scientist's Toolkit: Essential Research Reagents
The following tools and datasets are essential for conducting rigorous evaluations of structural prediction models.
Table 3: Key Resources for Benchmarking Biomolecular Predictions
| Tool / Dataset | Type | Primary Function in Evaluation |
|---|---|---|
| PoseBusters [4] | Software & Benchmark Dataset | Validates predicted protein-ligand complexes for steric clashes, bond geometry, and other physico-chemical plausibility metrics. |
| DockQ [7] [25] | Software & Metric | Provides a single continuous score for evaluating the quality of protein-protein and antibody-antigen docking models. |
| SAbDab [7] | Database | The primary repository for antibody and nanobody structural data, used for curating benchmark sets. |
| Gnina [4] | Software (CNN Scorer) | A deep learning-based scoring function used to re-rank docking poses, improving selection accuracy. |
| RDKit | Software (Cheminformatics) | A foundational toolkit for generating valid, diverse ligand conformations for docking inputs. |
| AlphaFold Server | Web Service | The primary interface for running non-commercial predictions with AlphaFold 3. |
The evidence indicates that there is no single superior tool for all scenarios; rather, the choice depends on the research question and available information. The following workflow can help researchers select the appropriate tool.
In summary, while AlphaFold 3 represents a transformative leap in the holistic prediction of biomolecular complexes, its reliance on pattern learning can sometimes lead to a compromise on strict physical realism, manifesting as steric clashes in challenging scenarios [6]. Physics-based docking methods, especially when enhanced with machine learning scoring and proper conformational sampling, remain robust and highly accurate alternatives, particularly when an experimental protein structure is available and the focus is on drug-like molecules [4]. For the foreseeable future, a synergistic approach—using AF3 for blind complex prediction and robust docking baselines for refinement and specific protein-ligand applications—will be the most reliable strategy for computational researchers. All computational predictions, regardless of the tool, should be considered hypotheses until validated by experimental data.
The accurate prediction of protein-ligand complex structures is a cornerstone of computational drug discovery. While the advent of deep learning systems like AlphaFold 3 (AF3) has revolutionized structural biology, their performance on biomolecular interactions with unseen scaffolds or novel targets remains a critical benchmarking frontier. This guide objectively compares the generalization capabilities of AF3 against established molecular docking methods, drawing on recently published data and benchmarks to inform researchers and development professionals.
Generalization—the ability of a model to make accurate predictions on inputs distinct from its training data—is particularly crucial in drug discovery, where researchers frequently investigate novel chemical matter against protein targets with limited structural characterization. This evaluation focuses specifically on performance with unseen ligand scaffolds and novel protein binding pockets, scenarios that closely mimic real-world drug discovery challenges.
Performance Comparison: Quantitative Benchmarks
Independent studies have evaluated AF3 and various docking approaches across multiple benchmark datasets designed to test generalization. The results reveal distinct performance patterns across different challenge levels.
Table 1: Overall Performance on Generalization Benchmarks
| Method | Type | Astex Diverse Set (RMSD ≤2Å & PB-valid) | PoseBusters Benchmark (RMSD ≤2Å & PB-valid) | DockGen (Novel Pockets) |
|---|---|---|---|---|
| AlphaFold 3 | Co-folding DL | Data not fully quantified | ~50% (blind), ~70% (pocket-specified) [4] | Performance decline reported [37] |
| Glide SP | Traditional Docking | >90% [37] | >90% [37] | >90% [37] |
| SurfDock | Generative Diffusion | 61.2% [37] | 39.3% [37] | 33.3% [37] |
| Strong Baseline (Vina + Gnina) | Hybrid Docking | Not tested | 69.2% (outperforms blind AF3) [4] | Not tested |
The data reveals a clear performance hierarchy, with traditional docking methods like Glide SP maintaining high success rates across all datasets, while deep learning methods, including AF3 and generative diffusion models, show more significant performance declines on novel pockets [37]. A specifically engineered strong baseline using Vina with Gnina rescoring and conformational ensembles demonstrated 69.2% success on the PoseBusters benchmark, outperforming the blind version of AF3 and approaching the accuracy of AF3 with specified pocket information [4].
Table 2: Performance on Different Ligand Types
| Method | Common Natural Ligands | Other Molecules (More Drug-like) |
|---|---|---|
| AlphaFold 3 | Excels (High accuracy) [4] | Lower performance [4] |
| Strong Baseline (Vina + Gnina) | Lower performance [4] | 8.5% above AF3 [4] |
AF3 demonstrates exceptional performance on common natural ligands (e.g., nucleotides, nucleosides) that are well-represented in its training data but shows relatively weaker performance on other, more drug-like molecules [4]. This suggests that the chemical space of typical small-molecule therapeutics may represent a generalization challenge for AF3.
Methodologies: Experimental Protocols for Evaluating Generalization
The PoseBusters Benchmark Protocol
The PoseBusters benchmark, used in the AF3 paper and subsequent independent evaluations, provides a standardized methodology for assessing prediction quality beyond simple RMSD metrics [4] [3] [37].
- Dataset Curation: Comprises protein-ligand structures released to the PDB after AlphaFold's training data cutoff (2021 or later) [4] [3]. This ensures the benchmark tests generalization to truly unseen structures.
- Validation Metrics: Evaluates predictions using both:
- Testing Modes:
- Blind Docking: Only protein sequence and ligand SMILES string are provided.
- Pocket-Specified Docking: Protein residues near the ligand are identified, providing additional spatial constraints [4].
Adversarial Testing for Physical Understanding
Recent research has employed adversarial examples based on physical principles to stress-test the generalization of co-folding models like AF3 [6].
- Binding Site Mutagenesis: Residues in the binding site are systematically mutated to disrupt favorable interactions.
- Glycine Scan: All binding site residues mutated to glycine, removing side-chain interactions.
- Phenylalanine Scan: All binding site residues mutated to phenylalanine, sterically occluding the pocket.
- Dissimilar Residue Mutation: Residues mutated to chemically dissimilar amino acids [6].
- Evaluation: Measures whether the model correctly predicts ligand displacement upon introducing disruptive mutations, testing if predictions are based on physical principles versus pattern matching to training data [6].
Cross-Docking and Apo-Docking Benchmarks
These protocols specifically test generalization to novel protein conformational states [1].
- Cross-Docking: Docking ligands to receptor conformations derived from different ligand complexes.
- Apo-Docking: Using unbound (apo) receptor structures from crystal structures or computational predictions.
- Significance: These scenarios require models to account for protein flexibility and induced fit effects, mimicking real-world drug discovery where experimental holo structures are often unavailable [1].
Visualization of Experimental Workflows
Adversarial Binding Site Mutagenesis
Performance Evaluation Workflow
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Software Tools for Docking Evaluation
| Tool | Type | Primary Function | Application in Generalization Testing |
|---|---|---|---|
| PoseBusters [4] [37] | Validation software | Automated quality checks for predicted structures | Detects steric clashes, stereochemical errors, and other physical implausibilities |
| Gnina [4] | Deep learning scoring function | Rescoring docked poses using neural networks | Improves pose selection in docking workflows |
| RDKit [4] | Cheminformatics toolkit | Generates ligand conformational ensembles | Enhances sampling for small molecule docking |
| AutoDock Vina [4] [38] | Molecular docking engine | Search-and-score based docking | Baseline method; component of strong docking pipelines |
| DiffDock [1] [37] | Deep learning docking | Generative diffusion model for blind docking | State-of-the-art DL method for comparison studies |
The generalization challenge represents a significant frontier in protein-ligand structure prediction. Current evidence suggests that while AF3 achieves remarkable accuracy on biomolecular complexes similar to its training data, its performance can decline on novel targets, particularly for drug-like small molecules and proteins with binding pockets distinct from those in the structural database [4] [37].
Physical adversarial tests reveal that co-folding models may sometimes prioritize pattern recognition over physical principles, continuing to place ligands in mutated binding sites that should no longer accommodate them [6]. This indicates potential limitations in their ability to generalize based on fundamental physics.
For researchers investigating novel targets or designing new chemical scaffolds, hybrid approaches that combine deep learning with physics-based methods may offer the most robust solution. Integrating AF3's pattern recognition strengths with the physical fidelity and proven generalization of traditional docking methods represents a promising direction for future methodological development.
In the field of computational structural biology, confidence metrics are indispensable for assessing the reliability of predicted models, guiding their application in downstream research, and interpreting results with appropriate caution. For protein structure prediction tools like AlphaFold 3 (AF3), two primary metrics—pLDDT (predicted local distance difference test) and pTM (predicted template modeling score)—provide complementary views of model quality. These metrics are particularly crucial when comparing the performance of deep learning-based co-folding models like AF3 against traditional molecular docking methods for predicting protein-ligand complexes, often referred to as "pose prediction" research.
Understanding these metrics allows researchers to gauge which regions of a predicted structure can be trusted for functional interpretation, drug binding site analysis, or rational protein engineering. This guide provides a comprehensive comparison of how these metrics are used to evaluate AF3's performance against specialized docking tools, complete with experimental data and methodologies to inform research decisions.
Understanding pLDDT and pTM
pLDDT: Local Per-Residue Confidence
The pLDDT is a per-residue measure of local confidence in a predicted structure, scaled from 0 to 100 [39]. It estimates how well the prediction would agree with an experimental structure using the local distance difference test (lDDT-Cα), a superposition-free metric that assesses the correctness of local distances [39] [40].
The pLDDT score is interpreted through established confidence bands:
- pLDDT > 90: Very high confidence; both backbone and side chains are typically predicted with high accuracy, with χ1 rotamers approximately 80% correct
- 70 < pLDDT < 90: Confident; generally correct backbone prediction with potential side chain misplacement
- 50 < pLDDT < 70: Low confidence; the prediction should be interpreted with caution
- pLDDT < 50: Very low confidence; likely indicating intrinsically disordered regions or regions with insufficient evolutionary information [39] [40]
pLDDT can vary significantly along a protein chain, allowing users to identify which regions are reliably predicted versus those that are unstructured or lack sufficient data for confident prediction [39].
pTM and ipTM: Global and Interface Confidence
For complexes and multimers, AlphaFold 3 provides two additional key metrics:
- pTM: Predicted template modeling score estimates the global reliability of the entire protein complex structure
- ipTM: Interface predicted template modeling score specifically evaluates the confidence in interactions between protein subunits [41]
These metrics address a critical limitation of pLDDT, which measures only local confidence and does not reflect confidence in the relative positions or orientations of domains in a protein or subunits in a complex [39]. The ipTM is particularly valuable for assessing the reliability of predicted protein-protein interfaces in multimers.
Table 1: Key Confidence Metrics in AlphaFold 3
| Metric | Scale | Interpretation | Application Scope |
|---|---|---|---|
| pLDDT | 0-100 | Local residue-level accuracy | Per-residue reliability |
| pTM | 0-1 | Global complex structure quality | Overall model confidence |
| ipTM | 0-1 | Subunit interaction accuracy | Interface reliability |
AlphaFold 3 vs. Molecular Docking: Performance Comparison
Benchmarking Results
When benchmarked against specialized molecular docking tools, AlphaFold 3 demonstrates remarkable performance in protein-ligand pose prediction, though important caveats exist regarding its physical understanding.
Table 2: Performance Comparison in Protein-Ligand Pose Prediction
| Method | Category | Accuracy (Ligand RMSD < 2Å) | Key Characteristics |
|---|---|---|---|
| AlphaFold 3 | Co-folding DL | 81% (blind), 93% (with site) | End-to-end complex prediction |
| DiffDock | Specialized DL | 38% (blind docking) | Deep learning docking |
| AutoDock Vina | Physics-based docking | ~60% (with known site) | Traditional scoring functions |
| RoseTTAFold All-Atom | Co-folding DL | Lower than AF3 | Similar approach to AF3 |
According to evaluations on the PoseBusterV2 dataset, AF3 achieved approximately 81% accuracy for blind docking (predicting native pose within 2Å RMSD) compared to DiffDock's 38% [6]. When the binding site is provided, AF3's accuracy exceeds 93%, significantly outperforming traditional physics-based docking methods like AutoDock Vina, which achieves approximately 60% accuracy under similar conditions [6].
Advantages of AlphaFold 3's Approach
AF3's architecture represents a substantial evolution from previous versions, contributing to its enhanced performance:
- Unified Framework: AF3 uses a single model to predict complexes containing proteins, nucleic acids, small molecules, ions, and modified residues, unlike specialized docking tools focused on specific interaction types [3]
- Diffusion-Based Architecture: Replaces AlphaFold 2's structure module with a diffusion module that directly predicts raw atom coordinates, eliminating the need for amino-acid-specific frames and torsion angles [3] [8]
- Reduced MSA Dependence: Incorporates a simpler "Pairformer" module that de-emphasizes multiple sequence alignment processing compared to AF2, potentially improving performance on targets with limited evolutionary data [41] [3]
This architecture allows AF3 to natively model conformational changes during binding, a significant challenge for traditional docking approaches that often treat proteins as rigid bodies [8].
Experimental Protocols for Validation
Binding Site Mutagenesis Challenge
Recent research has employed adversarial testing to evaluate whether deep learning models like AF3 truly learn the physics of molecular interactions or primarily rely on pattern recognition from training data.
Objective: To assess if co-folding models understand physical principles by testing predictions under biologically implausible binding site conditions [6].
Methodology:
- Select a protein-ligand complex with known structure (e.g., ATP binding to CDK2)
- Systematically mutate all binding site residues to:
- Glycine (removing side-chain interactions)
- Phenylalanine (sterically occluding the binding pocket)
- Dissimilar residues (drastically altering chemical properties)
- Compare predicted ligand poses against expected physical behavior
Key Findings: In glycine mutagenesis, all co-folding models (including AF3, RFAA, Chai-1, Boltz-1) continued predicting ATP binding despite loss of anchoring interactions. In phenylalanine challenges, predictions remained biased toward original binding sites, with some instances of unphysical atomic clashes [6].
Cross-Docking Benchmark Protocols
Standardized benchmarks are essential for fair comparison between AF3 and docking methods.
Dataset Preparation:
- Use the PoseBuster benchmark (428 protein-ligand structures released to PDB in 2021 or later) to ensure temporal separation from training data [3]
- For docking comparisons, use the CASF-2016 benchmark with 285 protein-ligand PDB structures organized around 57 targets [42]
Evaluation Metrics:
- Ligand RMSD: Measure root-mean-square deviation of predicted ligand pose after aligning protein binding sites
- Success Rate: Calculate percentage of predictions with ligand RMSD < 2Å
- Physical Plausibility: Check for steric clashes, improper bond lengths, and violation of physical constraints
Implementation Details:
- For AF3: Input protein sequence and ligand SMILES without structural information
- For docking tools: Use native protein structures for fair comparison
- For blind docking: Provide only protein sequence/structure without binding site information
- For site-specific docking: Provide binding site coordinates [6]
Critical Limitations and Physical Understanding
Despite impressive benchmark performance, critical studies question whether AF3 and similar co-folding models genuinely learn physical principles or primarily excel at pattern recognition from training data.
Robustness to Physically Implausible Perturbations
Recent adversarial testing reveals significant limitations in AF3's physical understanding. When binding site residues were mutated to glycine (removing side-chain interactions) or phenylalanine (sterically blocking the pocket), AF3 and other co-folding models continued predicting ligand binding in the original location, despite the absence of favorable interactions or presence of steric hindrance [6].
These findings indicate that rather than learning fundamental physics, these models may be overfitting to statistical correlations in their training data, potentially limiting generalization to novel protein-ligand systems not represented in the training distribution [6].
Comparison to Physics-Based Docking
Traditional docking methods employ explicit physical scoring functions with different strengths and limitations:
Table 3: Scoring Function Categories in Molecular Docking
| Scoring Type | Basis | Advantages | Limitations |
|---|---|---|---|
| Physics-Based | Force fields, molecular mechanics | Explicit physical basis | Computationally expensive, approximations |
| Empirical-Based | Weighted energy terms | Faster computation, simpler | Parameterization dependent |
| Knowledge-Based | Statistical potentials from known structures | Balance of speed and accuracy | Limited by database coverage |
| ML/DL-Based | Learned patterns from data | Can capture complex relationships | Black box, data-dependent |
While AF3 significantly outperforms these methods in benchmark accuracy, its occasional failure to respect basic physical principles suggests that traditional docking with explicit physical scoring may still offer advantages for certain applications requiring strict physical plausibility [6] [43].
The Scientist's Toolkit: Essential Research Reagents
Table 4: Key Resources for Pose Prediction Research
| Resource | Type | Function | Access |
|---|---|---|---|
| AlphaFold Server | Web Server | AF3 predictions with confidence metrics | https://alphafoldserver.com/ |
| PoseBusterV2 Dataset | Benchmark Dataset | Protein-ligand structures for validation | [6] |
| CASF-2016 | Benchmark Dataset | Standard set for scoring function comparison | [42] |
| CCharPPI Server | Evaluation Tool | Scoring function assessment independent of docking | [43] |
| Ligand B-Factor Index (LBI) | Quality Metric | Prioritizes complexes based on ligand vs. binding site flexibility | https://chembioinf.ro/tool-bi-computing.html [42] |
| PDB | Database | Experimental structures for validation/templates | https://www.rcsb.org/ |
Experimental Workflow Visualization
Diagram Title: Pose Prediction Research Workflow
Confidence metrics pLDDT and pTM are essential tools for assessing the reliability of AlphaFold 3 predictions in pose prediction research. While AF3 demonstrates remarkable accuracy in benchmark comparisons against specialized docking tools, researchers should interpret its predictions with awareness of its limitations in physical understanding.
For critical applications in drug discovery and protein engineering, a hybrid approach that leverages AF3's pattern recognition capabilities while validating against physical principles may offer the most robust strategy. The ongoing development of adversarial testing methodologies and more physically-grounded benchmarks will further enhance our ability to gauge the true reliability of these transformative deep learning tools.
AlphaFold 3 (AF3) represents a transformative advancement in biomolecular structure prediction, demonstrating exceptional accuracy in predicting protein-ligand complexes. Independent validation during CASP16 revealed that AF3 achieved a mean LDDT-PLI score of 0.8, outperforming the best human predictor group and establishing a new benchmark for computational pose prediction [44]. This performance is particularly notable in direct comparison experiments, where AF3 demonstrated approximately 81% accuracy in blind docking of small molecules compared to 38% for DiffDock, and over 93% accuracy when binding sites were provided compared to about 60% for AutoDock Vina [6].
However, despite these impressive capabilities, critical limitations persist that hinder AF3's standalone reliability for drug discovery applications. Recent investigations reveal that AF3 and similar co-folding models exhibit significant deviations from fundamental physical principles when subjected to biologically plausible perturbations [6]. In binding site mutagenesis experiments, these models continued to place ligands in original binding sites even after removing all favorable interactions, indicating potential overfitting to statistical patterns rather than learning underlying physics [6]. Furthermore, AF3 produces static structural snapshots that cannot capture dynamic conformational changes, lacks binding affinity predictions essential for drug development, and demonstrates limited generalization to novel protein binding pockets and specific challenges like modeling PROTAC ternary complexes [10] [45].
These limitations have catalyzed the development of hybrid strategies that integrate AF3's exceptional initial pose prediction with physics-based refinement to produce more biologically realistic and therapeutically relevant models.
Performance Comparison: AF3 vs. Traditional Docking Methods
Quantitative Accuracy Assessment
Table 1: Comparative Pose Prediction Accuracy Across Methodologies
| Method Category | Representative Tools | Pose Accuracy (RMSD ≤ 2Å) | Physical Validity (PB-valid Rate) | Combined Success (RMSD ≤ 2Å & PB-valid) | Key Strengths | Key Limitations |
|---|---|---|---|---|---|---|
| Co-folding Models | AlphaFold 3 | 77-94% [6] | Not reported | Not reported | Holistic complex modeling; Superior blind docking | Limited physical robustness; No affinity scores |
| Generative Diffusion | SurfDock, DiffBindFR | 70-92% [37] | 40-64% [37] | 33-61% [37] | Excellent pose accuracy | Moderate physical validity; Steric clashes |
| Traditional Methods | Glide SP, AutoDock Vina | Moderate (specific values not reported) [37] | 94-98% [37] | Moderate (specific values not reported) [37] | Excellent physical plausibility | Computationally intensive; Search limitations |
| Regression-based DL | KarmaDock, QuickBind | Low to moderate [37] | Low [37] | Low [37] | Fast predictions | Frequently invalid physical poses |
| Hybrid Methods | Interformer | Moderate [37] | High [37] | Superior balance [37] | Balanced accuracy & physicality | Implementation complexity |
Specialized Application Performance
In antibody-antigen docking, AF3 achieves a 10.2% high-accuracy docking success rate (DockQ ≥ 0.80) with single seed sampling, significantly outperforming AF2.3-Multimer's 2.4% success rate [7]. However, this still leaves a 65% failure rate for antibody and nanobody docking, indicating substantial room for improvement [7].
For complex applications like PROTAC ternary complexes, AF3's performance appears inflated by accessory proteins that contribute to interface area but not degrader-specific binding. When evaluated on core complex components, PRosettaC, which leverages chemically defined anchor points, outperforms AF3 in geometric accuracy [45].
Experimental Protocols for Hybrid Workflow Development
Integrated AF3-Physics Refinement Pipeline
The hybrid methodology emerges from systematic evaluations demonstrating complementary strengths: AF3 provides superior initial pose generation, while physics-based methods ensure physical plausibility and refinement.
Table 2: Core Experimental Methodologies for Validation
| Methodology | Experimental Purpose | Key Metrics | Implementation Tools |
|---|---|---|---|
| PoseBusters Validation [37] | Assess physical plausibility of predictions | Bond lengths/angles, stereochemistry, steric clashes | PoseBusters toolkit |
| Binding Site Mutagenesis [6] | Test model robustness & physical understanding | Ligand displacement response, steric clashes | Residue substitution scans |
| Molecular Dynamics (MD) Simulations [14] [45] | Evaluate structural stability & conformational sampling | RMSD evolution, intermolecular interactions, energy profiles | GROMACS, AMBER, OpenMM |
| Frame-Resolved DockQ Analysis [45] | Dynamic assessment of interface quality | DockQ scores across MD trajectories | Custom analysis scripts |
| Alanine Scanning [14] | Identify critical binding residues | Binding affinity changes (ΔΔG) | MM-GBSA, MM-PBSA |
Reference Experimental Workflows
Protocol 1: Basic AF3 Pose Refinement
- AF3 Prediction: Generate initial complexes using AF3 Server with default parameters [10]
- Confidence Assessment: Identify high-confidence regions using pLDDT and ipTM scores [10] [7]
- Physics-Based Minimization: Apply force field-based relaxation (AMBER, CHARMM) to resolve steric clashes
- Explicit Solvent MD: Run 10-100ns simulations to sample flexible regions [14]
- Cluster Analysis: Extract representative poses from MD trajectories for binding assessment
Protocol 2: Virtual Screening Hybrid Approach
- Binding Site Identification: Use AF3 for blind pocket detection [1]
- Multi-Conformer Sampling: Generate diverse ligand poses using AF3 sampling [10]
- Physics-Based Scoring: Re-rank poses using MM-GBSA or FEP calculations [14]
- Ensemble Docking: Employ multiple AF3-generated receptor conformations [1]
- Experimental Validation: Verify top candidates through crystallography or functional assays [44]
Implementation Framework for Hybrid Methodologies
Workflow Architecture
The following diagram illustrates the integrated hybrid workflow that combines AF3's sampling capabilities with physics-based validation and refinement:
Essential Research Reagents and Computational Tools
Table 3: Research Reagent Solutions for Hybrid Method Implementation
| Category | Tool/Resource | Function | Access Considerations |
|---|---|---|---|
| Structure Prediction | AlphaFold Server | Initial complex prediction | Free academic access; Non-commercial use only [10] |
| Physical Validation | PoseBusters Toolkit | Geometric and steric validation | Open source [37] |
| Molecular Dynamics | GROMACS, AMBER, OpenMM | Physics-based sampling and refinement | Open source or academic licensing |
| Scoring Functions | MM-GBSA, MM-PBSA | Binding affinity estimation | Built into MD packages |
| Specialized Docking | PRosettaC | PROTAC ternary complex modeling | Open source [45] |
| Confidence Metrics | pLDDT, ipTM | Prediction reliability assessment | AF3 server output [10] [7] |
Hybrid strategies that integrate AlphaFold 3's exceptional pattern recognition with physics-based refinement represent a paradigm shift in computational structural biology. The experimental evidence consistently demonstrates that while AF3 provides unprecedented initial pose accuracy, its integration with physical principles addresses critical limitations in modeling dynamic behavior, physical plausibility, and binding energetics.
Future developments will likely focus on tightly integrated pipelines that seamlessly combine deep learning and physics-based approaches, dynamic sampling techniques that go beyond static snapshots, and specialized applications for challenging targets like membrane proteins and flexible systems. As the field evolves, the most successful implementations will be those that leverage the complementary strengths of data-driven prediction and first-principles physics, ultimately accelerating drug discovery through more reliable in silico structural modeling.
The benchmark data and methodologies presented provide researchers with a framework for developing and validating these hybrid approaches, emphasizing the importance of physical validation and experimental correlation to ensure predictive reliability in real-world drug discovery applications.
Critical Validation: Stress-Testing Against Physical and Biological Principles
The accurate prediction of protein-ligand complexes represents a cornerstone of modern computational biology, with profound implications for drug discovery and protein engineering. Two dominant paradigms have emerged in this field: traditional molecular docking tools, which rely on physics-based scoring functions and sampling algorithms, and the newer deep learning-based co-folding models, such as AlphaFold 3 (AF3), which use end-to-end neural networks to predict complex structures directly from sequence and chemical information [6] [3]. While benchmarks often show co-folding models achieving superior accuracy on standard test sets, their reliance on pattern recognition from training data raises critical questions about their true understanding of underlying physical principles [6] [14].
This guide objectively compares the performance of AF3 and other co-folding models against the backdrop of traditional docking, specifically under adversarial testing conditions. Adversarial tests, such as binding site mutagenesis and ligand perturbation, probe model robustness by introducing biologically plausible but challenging modifications that disrupt native interactions [6]. Such tests move beyond standard benchmarks to reveal whether models are learning the fundamental physics of molecular interactions or merely memorizing statistical correlations present in their training data. The findings summarized here provide crucial insights for researchers relying on these tools for critical applications in drug discovery and protein design.
Performance Comparison Under Adversarial Conditions
Binding Site Mutagenesis Challenges
A pivotal study investigated the robustness of deep-learning co-folding models by subjecting them to a series of binding site mutagenesis challenges on the Cyclin-dependent kinase 2 (CDK2) protein in complex with its native ligand, ATP [6]. The models were tasked with predicting the structure of the complex after the binding site residues were systematically mutated in ways that should, based on biophysical principles, displace the ligand.
Table 1: Performance on Binding Site Mutagenesis Challenges (CDK2-ATP Complex)
| Adversarial Challenge | Description | AlphaFold 3 | RoseTTAFold All-Atom | Chai-1 | Boltz-1 |
|---|---|---|---|---|---|
| Wild-Type (No Mutation) | Baseline prediction against native crystal structure. | RMSD: 0.2 Å (High Accuracy) | RMSD: 2.2 Å | High Accuracy | High Accuracy |
| Glycine Scan | All binding site residues replaced with glycine. | Loses precise placement, but ligand remains in site. | Ligand remains (RMSD: 2.0 Å). Few/no interactions. | Ligand pose mostly unchanged. | Slight change in triphosphate position. |
| Phenylalanine Scan | All binding site residues replaced with phenylalanine. | Ligand pose biased to original site; minor adjustments. | Ligand entirely within original site; steric clashes. | Ligand entirely within original site. | Ligand pose biased to original site. |
| Dissimilar Residue Mutation | Residues mutated to alter shape/chemistry. | No significant pose alteration; significant steric clashes. | No significant pose alteration; significant steric clashes. | No significant pose alteration. | No significant pose alteration. |
*Table 1 summarizes the performance of four co-folding models when the binding site of CDK2 is adversarially mutated. RMSD (Root Mean Square Deviation) measures the difference in ligand position between the prediction and the experimental structure. A lower RMSD indicates a more accurate prediction. The results reveal a common limitation: despite the radical removal of favorable interactions and the introduction of steric hindrance, these models display a strong prediction bias towards the original, native binding pose [6]. This suggests that for well-characterized systems like ATP-binding proteins, the models may be overfitting to patterns in the training data rather than inferring the functional consequences of the introduced mutations.
Comparison with Traditional Docking
Traditional docking tools like AutoDock Vina operate on a different principle. They perform a conformational search for the ligand within a defined binding site, guided by a physics-inspired scoring function [6] [46]. While their performance can degrade if the binding site conformation is incorrect, they are inherently responsive to changes in the protein's atomic structure because their scoring function explicitly calculates interactions based on the provided atomic coordinates.
The key difference illuminated by adversarial testing is that docking algorithms explicitly compute interactions for the given protein structure, whereas co-folding models appear to implicitly predict them based on learned sequence-structure relationships. Consequently, docking tools would be expected to correctly predict ligand displacement in the aforementioned mutagenesis challenges, as their scoring function would no longer favor the mutated binding site.
Experimental Protocols for Adversarial Testing
The following section outlines the methodologies used in the key studies cited in this guide, providing a protocol for researchers seeking to perform similar robustness evaluations.
Protocol: Binding Site Mutagenesis Challenge
This protocol is derived from the study that tested AF3 and other models on mutated CDK2 [6].
- System Selection: Select a high-quality protein-ligand complex structure from a database like the PDB (e.g., CDK2 with ATP). The complex should have a well-defined binding site.
- Baseline Prediction: Input the wild-type protein sequence and ligand description (e.g., SMILES string) into the co-folding model (AF3 server, etc.) and/or docking software to establish baseline prediction accuracy.
- Define Binding Site Residues: Identify all residues with atoms within a specified distance (e.g., 4-5 Å) of the bound ligand.
- Design Mutations: Create a series of mutant protein sequences:
- Glycine Scan: Mutate all binding site residues to glycine. This removes side-chain interactions and increases pocket flexibility.
- Phenylalanine Scan: Mutate all binding site residues to phenylalanine. This removes favorable chemical interactions and sterically occludes the binding pocket with bulky aromatic rings.
- Dissimilar Residue Mutation: Mutate each binding site residue to a chemically and structurally dissimilar amino acid (e.g., polar to hydrophobic, small to bulky).
- Run Predictions: Submit each mutant sequence along with the same ligand description to the prediction models.
- Analysis: Compare the predicted ligand pose for each mutant to the wild-type prediction and the original crystal structure. Key metrics include:
- Ligand RMSD.
- Presence of steric clashes.
- Loss of specific interactions (e.g., hydrogen bonds, electrostatic interactions).
Protocol: Leveraging Experimental Data for Docking
A separate methodology highlights how traditional docking can be enhanced by integrating experimental data, a flexibility not currently available in closed co-folding systems like AF3 [46].
- Obtain Experimental Density: Acquire the underlying electron density map from a cryo-EM or X-ray crystallography experiment.
- Process Density: Use a tool like CryoXKit to convert the experimental density into a biasing potential for docking [46].
- Perform Guided Docking: Run the docking simulation (e.g., using AutoDock-GPU) with the added biasing potential, which guides ligand heavy atoms towards regions of high electron density.
- Analysis: This approach has been shown to significantly improve pose prediction in both re-docking and cross-docking scenarios, and can enhance virtual screening performance [46].
Research Reagent Solutions
The table below catalogues key computational tools and resources mentioned in this guide that are essential for conducting research in protein-ligand structure prediction and adversarial testing.
Table 2: Key Research Reagents and Tools
| Tool / Resource | Type | Primary Function | Relevance to Adversarial Testing |
|---|---|---|---|
| AlphaFold Server | Web Server | Free academic platform for predicting structures of protein complexes with ligands, nucleic acids, and more using AF3 [10]. | Primary tool for testing AF3's performance on wild-type and adversarially modified sequences. |
| RoseTTAFold All-Atom (RFAA) | Software Tool | An open-source deep learning model for predicting structures of biomolecular complexes, similar to AF3 [6]. | An alternative co-folding model for comparative robustness analysis. |
| AutoDock Vina/GPU | Software Tool | A widely used, physics-based molecular docking program for predicting protein-ligand binding poses and scoring [6] [46]. | Represents the traditional docking paradigm; responsive to explicit atomic changes. |
| CryoXKit | Software Tool | A tool that processes cryo-EM or X-ray crystallography density maps to create a biasing potential for docking [46]. | Enhances docking accuracy by incorporating experimental data, a hybrid approach. |
| Boltz-2 | Software Tool | An open-source model that predicts both protein-ligand complex structure and binding affinity [47]. | Represents the next generation of models that go beyond structure to functional properties. |
| Protein Data Bank (PDB) | Database | A repository for the 3D structural data of large biological molecules [3]. | Source for obtaining wild-type protein-ligand complex structures to establish ground truth. |
The following diagram illustrates the logical workflow and core findings of the binding site mutagenesis study, highlighting the divergent behaviors of co-folding models and traditional docking tools.
Adversarial testing through binding site mutagenesis provides a necessary and revealing stress test for protein-ligand structure prediction tools. The experimental data demonstrates that while deep learning co-folding models like AlphaFold 3 achieve stunning accuracy on standard benchmarks, they can fail to generalize when presented with biologically plausible but adversarial inputs [6]. Their predictions often remain stubbornly biased toward the native binding mode, even after removing key interacting residues, indicating a potential over-reliance on statistical patterns in the training data rather than a robust understanding of physical chemistry.
In contrast, traditional molecular docking methods, while often less accurate on standard tests and reliant on a predefined binding site, are inherently more responsive to atomic-level changes in the protein because they explicitly compute interactions for the provided structure. The choice between these paradigms, therefore, depends on the research context. For predicting structures of wild-type complexes, AF3 is a powerful and often superior tool. However, for applications involving mutated proteins, drug design for novel binding sites, or any scenario requiring a deep understanding of physicochemical principles, traditional docking or the emerging hybrid approaches that integrate experimental data [46] and physical models [47] remain essential. A measured, complementary use of both classes of tools, with a clear awareness of their respective strengths and weaknesses, is the most prudent path forward for critical research in drug discovery and structural biology.
Molecular docking, a cornerstone of computational drug discovery, aims to predict the three-dimensional structure of protein-ligand complexes and their binding affinity. For decades, the majority of docking approaches treated proteins as rigid bodies while allowing varying degrees of ligand flexibility. This simplification significantly limited predictive accuracy because proteins are inherently dynamic entities that undergo conformational changes upon ligand binding—a phenomenon known as induced fit [1] [48]. The limitations of rigid receptor assumptions become particularly evident in two challenging docking scenarios: cross-docking and apo-docking.
Cross-docking involves docking ligands to alternative receptor conformations derived from different protein-ligand complexes, simulating real-world cases where ligands are docked to proteins in unknown conformational states [1]. Apo-docking uses unbound (apo) receptor structures, typically obtained from crystal structures or computational predictions, requiring models to infer the induced fit and accommodate structural differences between unbound and bound states [1]. These scenarios represent more realistic and challenging tasks compared to re-docking (docking a ligand back into its original receptor structure), where performance is typically much higher [49].
The emergence of deep learning approaches like AlphaFold 3 has revolutionized structural biology and molecular docking by offering unprecedented accuracy in predicting protein-ligand interactions [3]. This review provides a comprehensive comparison between traditional docking methods and AlphaFold 3, specifically evaluating their performance in handling receptor flexibility through cross-docking and apo-docking scenarios, with supporting experimental data and detailed methodological protocols.
Defining the Docking Challenges: Cross-Docking and Apo-Docking
Molecular docking tasks vary significantly in their complexity and constraints, primarily determined by the structural information provided about the receptor. The table below summarizes the key docking tasks relevant to flexibility assessment.
Table 1: Molecular Docking Tasks and Their Characteristics
| Docking Task | Description | Key Challenge | Real-World Relevance |
|---|---|---|---|
| Re-docking | Docking a ligand back into the bound (holo) conformation of its original receptor | Limited utility for novel compounds | Low; primarily for method validation |
| Cross-docking | Docking ligands to alternative receptor conformations from different ligand complexes | Handling conformational variation between different bound states | High; simulates docking to proteins with known ligands |
| Apo-docking | Docking to unbound (apo) receptor structures | Predicting induced fit changes from apo to holo states | Very high; most common real-world scenario |
| Blind docking | Predicting both binding site location and ligand pose | No prior knowledge of binding site | Moderate; useful for novel target exploration |
The fundamental challenge in both cross-docking and apo-docking stems from the conformational plasticity of protein binding sites. Proteins exist as ensembles of states, and ligand binding often stabilizes particular conformations from this ensemble [48]. The structural spectrum can range from minor side-chain rearrangements to substantial backbone movements and domain shifts [1] [49]. Traditional docking methods struggle with these conformational changes because their scoring functions are typically optimized for static structures, and exhaustively sampling protein flexibility is computationally prohibitive [49].
Traditional Docking Approaches to Receptor Flexibility
Historical Evolution and Methodological Categories
Traditional molecular docking methods primarily follow a search-and-score framework, exploring possible ligand poses and predicting optimal binding conformations based on scoring functions that estimate protein-ligand binding strength [1]. The evolution of handling flexibility in traditional docking can be categorized into several approaches:
- Soft Docking: Uses softened potentials that limit penalties for minor steric clashes, allowing some structural ambiguity [49]
- Side-Chain Flexibility: Allows rotation of side-chain torsional angles while keeping the backbone fixed, often using rotamer libraries [49]
- Multiple Receptor Conformations (MRC): Uses multiple static protein structures to represent different conformational states [49]
- Collective Degrees of Freedom: Uses normal modes or other collective variables to describe large-scale protein motions [49]
Among these, the MRC approach (also called ensemble docking) has been particularly popular due to its practical implementation and reasonable computational demands [49]. This method involves docking against multiple protein structures either sequentially or through specialized ensemble docking algorithms.
Performance Limitations of Traditional Methods
Traditional docking approaches show significant performance degradation when moving from re-docking to more realistic cross-docking and apo-docking scenarios. State-of-the-art docking algorithms predict an incorrect binding pose for about 50-70% of all ligands when only a single fixed receptor conformation is considered [49]. Even when the correct pose is obtained, lack of receptor flexibility often results in meaningless binding scores that don't correlate with experimental affinities [49].
The MRC approach demonstrates that using multiple receptor conformations can improve both pose prediction and virtual screening performance. In studies on aldose reductase, MRC docking showed a 40% improvement over 'hard' docking to a single conformation, successfully identifying novel low-micromolar inhibitors [49]. However, performance gains are system-dependent and limited by the quality and diversity of the available receptor conformations.
AlphaFold 3: A Paradigm Shift in Biomolecular Structure Prediction
Architectural Innovations
AlphaFold 3 represents a fundamental transformation in biomolecular structure prediction through several key architectural innovations:
- Unified Diffusion-Based Architecture: Replaces the complex structure module of AlphaFold 2 with a diffusion approach that operates directly on raw atom coordinates, enabling joint prediction of complexes containing proteins, nucleic acids, small molecules, ions, and modified residues [3]
- Reduced MSA Processing: Replaces the evoformer with a simpler pairformer module, de-emphasizing multiple sequence alignment processing [3]
- Multiscale Denoising: The diffusion process learns protein structure at various length scales—small noise levels refine local stereochemistry while high noise levels emphasize large-scale structure [3]
- Cross-Distillation: Uses structures predicted by AlphaFold-Multimer to reduce hallucination of compact structures in unstructured regions [3]
These innovations enable AlphaFold 3 to handle diverse biomolecules within a single framework while naturally accommodating the flexibility required for accurate complex prediction.
Performance Advantages in Standard Benchmarks
AlphaFold 3 demonstrates remarkable performance advantages over specialized traditional methods. On the PoseBusters benchmark (composed of 428 protein-ligand structures), AlphaFold 3 achieves far greater accuracy for protein-ligand interactions compared with state-of-the-art docking tools, even while operating as a true blind predictor without structural inputs [3]. The model is 50% more accurate than the best traditional methods on this benchmark, making it the first AI system to surpass physics-based tools for biomolecular structure prediction [50].
Table 2: Performance Comparison on PoseBusters Benchmark
| Method | Input Type | Success Rate (% <2Å) | Relative Performance |
|---|---|---|---|
| AlphaFold 3 | Sequence + SMILES | Highest reported | 50% more accurate than best traditional methods |
| Traditional Docking | Structure + SMILES | Lower than AF3 | Requires receptor structure as input |
| RoseTTAFold All-Atom | Sequence + SMILES | Significantly lower | Greatly outperformed by AF3 |
Comparative Performance in Flexible Receptor Docking
Cross-Docking and Apo-Docking Performance
While AlphaFold 3 excels in standard benchmarks, its performance in more challenging flexible docking scenarios reveals both strengths and limitations. In apo-docking scenarios, where ligands are docked to unbound receptor structures, AlphaFold 3 demonstrates a remarkable ability to predict induced fit conformational changes without explicit training on these transitions [1].
However, recent evaluations on protein-PFAS (per- and polyfluoroalkyl substances) complexes reveal important nuances in AlphaFold 3's generalization capabilities. When tested on a "Before Set" (structures likely seen during training), AlphaFold 3 achieved approximately 74.5% success rate in pocket-aligned ligand predictions. This performance dropped to approximately 55.8% on an "After Set" (unseen structures), indicating potential overfitting to training data [22].
The following diagram illustrates the conceptual workflow and challenges of cross-docking and apo-docking evaluations:
Hybrid Approaches and Performance Enhancement Strategies
Research indicates that hybrid approaches combining AlphaFold 3 with traditional docking methods can leverage the strengths of both. A study on protein-PFAS interactions found that using AlphaFold 3 for binding pocket identification followed by AutoDock Vina for interaction modeling improved prediction accuracy compared to either method alone [22]. This suggests that AlphaFold 3's pocket prediction capabilities are robust, while pose refinement may benefit from physics-based scoring.
Similarly, the Folding-Docking-Affinity (FDA) framework demonstrates how combining ColabFold for protein structure prediction, DiffDock for docking, and GIGN for affinity prediction achieves performance comparable to state-of-the-art docking-free methods in kinase-specific benchmarks [51]. Surprisingly, using ColabFold-generated apo-structures sometimes yielded improved affinity prediction performance compared to crystallized holo-structures, highlighting the potential of computational structural models in docking pipelines [51].
Table 3: Performance Comparison Across Docking Scenarios and Methods
| Method | Re-docking Performance | Cross-docking Performance | Apo-docking Performance | Computational Cost |
|---|---|---|---|---|
| Traditional Docking (Single Structure) | High (Optimized for this scenario) | Low to Moderate (50-70% failure rate) | Low (Struggles with induced fit) | Low to Moderate |
| Traditional Docking (MRC) | High | Moderate (Depends on ensemble diversity) | Moderate (Limited by apo structures in ensemble) | Moderate to High (Scales with ensemble size) |
| AlphaFold 3 | Very High | High (but potential overfitting concerns) | High (Can predict conformational changes) | Moderate (GPU-intensive) |
| Hybrid Approaches (AF3 + Traditional) | High | Highest Reported (Leverages strengths of both) | Highest Reported (Combined pocket and pose accuracy) | High (Multiple method execution) |
Experimental Protocols for Method Evaluation
Standard Evaluation Metrics and Protocols
Rigorous evaluation of docking performance for flexible receptors requires standardized metrics and protocols:
- Success Rate: Typically defined as the percentage of protein-ligand pairs with pocket-aligned ligand root mean square deviation (RMSD) of less than 2Å from experimental structures [3] [22]
- Pocket-Aligned RMSD: Calculated after aligning the predicted structure to the experimental structure based on binding pocket atoms only, providing a more relevant measure than global RMSD [3]
- Generalization Assessment: Splitting test sets into "Before" and "After" sets based on release dates to evaluate performance on structures potentially seen during training versus completely novel structures [22]
- Cross-docking Specific Protocols: Using receptor conformations from different ligand complexes than the one being docked [1]
- Apo-docking Specific Protocols: Using truly unbound receptor structures rather than holo structures with ligands removed [1]
Data Set Curation and Preparation
Proper dataset curation is essential for meaningful evaluation:
- Temporal Splitting: Ensuring test structures were released after training cut-off dates to prevent data leakage [22]
- Diversity Considerations: Including proteins with varying degrees of flexibility and different types of conformational changes [1]
- Complexity Gradation: Testing performance across different difficulty levels from re-docking to blind docking [1]
- Experimental Structure Verification: Using high-resolution crystal structures with clear electron density for benchmark creation [49]
The following experimental workflow illustrates a comprehensive evaluation protocol for flexible receptor docking:
Table 4: Key Research Resources for Flexible Receptor Docking Studies
| Resource Category | Specific Tools/Services | Primary Function | Relevance to Flexible Docking |
|---|---|---|---|
| Structure Prediction | AlphaFold Server, ColabFold | Protein structure generation from sequence | Provides apo structures for docking when experimental structures are unavailable |
| Traditional Docking | AutoDock Vina, DOCK, FlexX | Pose prediction and scoring | Baseline methods for comparison and hybrid approaches |
| Specialized Flexibility Tools | FlexE, FLIPDock, FITTED | Explicit handling of receptor flexibility | Representative specialized traditional approaches |
| Benchmark Datasets | PDBBind, PoseBusters, DUD-E | Standardized performance assessment | Enables fair comparison across methods |
| Analysis and Visualization | PyMOL, Chimera, RDKit | Structure analysis and result interpretation | Critical for qualitative assessment of predictions |
| Force Fields | AMBER, CHARMM | Molecular mechanics calculations | Used in structure preparation and refinement |
The evaluation of docking performance with flexible receptors through cross-docking and apo-docking scenarios reveals a rapidly evolving landscape where deep learning approaches like AlphaFold 3 are setting new standards for accuracy. However, traditional methods remain relevant, particularly when integrated into hybrid approaches that leverage the complementary strengths of physical and learning-based methods.
Key findings from current research indicate:
AlphaFold 3 demonstrates superior performance in standard docking benchmarks, outperforming even specialized traditional tools while operating as a true blind predictor [3] [50]
Generalization to unseen structures remains challenging for all methods, with performance drops of nearly 20% observed for AlphaFold 3 when moving from training-like to novel structures [22]
Hybrid approaches combining AlphaFold 3's pocket identification with traditional docking's pose refinement show promise for achieving state-of-the-art performance in flexible receptor scenarios [22] [51]
Explicit modeling of protein flexibility through methods like FlexPose and DynamicBind represents the next frontier in addressing conformational diversity beyond what static structures can provide [1]
As the field progresses, the integration of more sophisticated flexibility modeling, improved generalization to novel targets, and streamlined workflows combining the strengths of multiple approaches will likely define the next generation of docking tools for drug discovery.
The emergence of deep learning (DL) has catalyzed a paradigm shift in biomolecular structure prediction, extending beyond single proteins to complex multimolecular assemblies. AlphaFold 3 (AF3), RoseTTAFold All-Atom (RFAA), Boltz-1, Chai-1, and DiffDock represent the vanguard of this revolution, enabling researchers to predict the structure of protein-ligand complexes with unprecedented accuracy. These advancements hold particular significance for drug discovery and development, where understanding molecular interactions at atomic resolution is paramount. This guide provides an objective, data-driven comparison of these five prominent methods, focusing on their performance in protein-ligand pose prediction within the specific context of evaluating AF3's capabilities against molecular docking alternatives. We synthesize evidence from recent benchmarking studies to delineate the relative strengths, limitations, and optimal use cases for each tool, providing researchers with a practical framework for method selection based on empirical evidence rather than anecdotal performance.
Performance Metrics and Benchmarking Results
Benchmarking studies consistently reveal that DL co-folding methods generally outperform traditional docking algorithms, with AF3, Boltz-1, and Chai-1 demonstrating particularly strong performance across diverse datasets.
Table 1: Overall Performance Metrics on Primary Ligand Docking Tasks
| Method | Type | Astex Diverse (RMSD ≤ 2Å & PB-Valid) | DockGen-E (RMSD ≤ 2Å & PB-Valid) | PoseBusters Benchmark (RMSD ≤ 2Å & PB-Valid) | BCAPIN (Acceptable Quality) |
|---|---|---|---|---|---|
| AlphaFold 3 (AF3) | Co-folding | High (~90%+) | <75% | Moderate | ~85% |
| Boltz-1 | Co-folding | High | Moderate | Moderate | ~85% |
| Chai-1 | Co-folding | High | Moderate | Comparable to AF3 | ~85% |
| RFAA | Co-folding | Moderate | Low | Low | ~85% |
| DiffDock | Docking | Lower than co-folding | Low | Low | ~85% |
Note: Performance metrics are relative comparisons based on aggregated results from multiple benchmarks. Exact percentages vary by dataset and evaluation criteria [52] [53] [54].
On the protein-carbohydrate-specific BCAPIN dataset, all five methods achieved comparable results with approximately 85% success rates for producing structures of at least acceptable quality [52] [53]. However, a critical limitation observed across all models was declining predictive power with increasing carbohydrate polymer length [52].
Confidence Metrics and Chemical Specificity
Confidence metrics such as pLDDT (predicted Local Distance Difference Test) and interface pTM (ipTM) provide crucial indicators of prediction reliability, with significant variation observed across methods.
Table 2: Confidence Metric Correlations and Chemical Specificity
| Method | Correlation (r) DockQC to pLDDT | PLIF-WM (Astex Diverse) | PLIF-WM (DockGen-E) | MSA Dependence |
|---|---|---|---|---|
| AF3 | 0.59 | High | Moderate | High |
| Boltz-1 | 0.64 | High | Moderate | Moderate |
| Chai-1 | 0.73 | High | Moderate | Low |
| RFAA | 0.79 | Moderate | Low | Moderate |
| DiffDock | N/A | Lower than co-folding | Low | N/A |
PLIF-WM (Protein-Ligand Interaction Fingerprint Wasserstein Matching Score) measures chemical specificity in recapitulating native amino acid-specific interaction patterns [53] [54].
Notably, AF3 demonstrates concerning overconfidence in certain contexts, with relatively weak correlation (r=0.59) between its confidence scores and actual accuracy on protein-carbohydrate complexes [53]. RFAA shows the strongest correlation (r=0.79) between pLDDT and accuracy metrics in the same benchmark [53]. Chai-1 exhibits lower dependence on multiple sequence alignments (MSAs), maintaining strong performance even in single-sequence mode, likely due to its incorporation of ESM2 language model embeddings during training [54].
Performance on Challenging and Multi-Ligand Targets
Performance disparities become more pronounced on challenging targets, such as those with novel binding poses or multiple ligands.
Table 3: Performance on Complex Prediction Scenarios
| Method | Multi-Ligand Docking | Novel/Uncommon Pockets | Performance on Long Carbohydrates |
|---|---|---|---|
| AF3 | Struggles with balance | Challenged by novel poses | Declining performance |
| Boltz-1 | Struggles with balance | Moderate | Declining performance |
| Chai-1 | Struggles with balance | Better generalization than AF3 | Declining performance |
| RFAA | Low accuracy | Low accuracy | Declining performance |
| DiffDock | Low accuracy | Low accuracy | Declining performance |
DL methods universally struggle to balance structural accuracy with chemical specificity when predicting novel protein-ligand binding poses or multi-ligand targets [54]. All models show reduced performance on longer carbohydrate polymers, highlighting a shared limitation in modeling extended sugar structures [52].
Computational Requirements and Runtime
Practical implementation considerations reveal substantial differences in computational resource requirements and processing speed.
Table 4: Computational Efficiency and Resource Requirements
| Method | Average Runtime | Memory Usage | Accessibility |
|---|---|---|---|
| AF3 | High | High | Limited (server access) |
| Boltz-1 | Moderate | Moderate | Open source |
| Chai-1 | Moderate | Moderate | Proprietary |
| RFAA | High | High | Open source |
| DiffDock | Low | Low | Open source |
Note: Metrics are relative comparisons based on PoseBench evaluations. Exact runtime and memory usage depend on hardware configuration and target complexity [54].
DiffDock generally offers the most efficient computation, while AF3 and RFAA require more substantial computational resources [54]. Access to AF3 is currently limited through a server interface, while Boltz-1, RFAA, and DiffDock are available as open-source tools, and Chai-1 operates as a proprietary platform [54].
Experimental Protocols and Methodologies
Standardized Benchmarking Frameworks
Rigorous evaluation of molecular docking and co-folding methods requires standardized datasets and metrics to ensure fair comparison.
PoseBench Evaluation Framework
PoseBench provides the first comprehensive benchmark for broadly applicable protein-ligand docking, specifically designed to assess performance in real-world scenarios [54]:
- Apo-to-Holo Prediction: Evaluates methods using predicted (apo) protein structures rather than experimental structures, reflecting the common scenario where only the unbound structure is available.
- Multi-Ligand Docking: Incorporates complexes with multiple bound ligands, addressing a critical gap in previous benchmarks.
- Blind Docking: Assesses performance without prior knowledge of binding pockets.
- Key Metrics: Includes percentage of predictions with RMSD ≤ 2Å, chemical validity (PB-Valid), and the novel PLIF-WM score for chemical specificity.
BCAPIN Dataset for Protein-Carbohydrate Complexes
The Benchmark of CArbohydrate Protein INteractions (BCAPIN) provides a specialized test set for evaluating protein-sugar interactions [52]:
- Curation: Derived from the DIONYSUS database of experimental protein-glycan structures from the PDB.
- Filtering: Protein sequences clustered at 50% identity, excluding structures solved before September 2021 (training cutoff for latest models).
- Quality Control: Filtered using Real Space Correlation Coefficient (RSCC) ≥ 0.9 to ensure structural reliability.
- Composition: 20 high-quality structures including 9 monomer-binding, 3 dimer-binding, 5 polymer-binding, and 3 with nucleotide and saccharide ligands.
- Evaluation Metric: DockQC, a novel metric inspired by protein-protein docking evaluation.
Adversarial Testing for Physical Realism
Recent research has employed adversarial examples to test whether co-folding models learn underlying physical principles rather than merely memorizing training data patterns [55] [6].
Binding Site Mutagenesis Protocol:
- Target Selection: Use well-characterized complexes such as ATP-bound CDK2.
- Wild-Type Prediction: Generate baseline predictions for unmodified complexes.
- Progressive Mutagenesis:
- Binding Site Removal: Replace all binding site residues with glycine.
- Steric Occlusion: Mutate binding site residues to phenylalanine.
- Chemical Incompatibility: Mutate to dissimilar residues that alter shape and chemical properties.
- Evaluation: Assess whether models adjust predictions appropriately or maintain biased poses despite unfavorable interactions.
Findings from these adversarial tests reveal that co-folding models often produce physically unrealistic structures, displaying bias toward training data and insufficient responsiveness to binding site modifications that should displace ligands [55] [6].
Workflow Visualization
The following diagram illustrates the comprehensive evaluation workflow used to benchmark these protein-ligand complex prediction methods, integrating both standard and adversarial testing protocols:
Diagram Title: Protein-Ligand Prediction Evaluation Workflow
Table 5: Key Experimental Resources and Their Applications
| Resource | Type | Primary Function | Relevance to Method Evaluation |
|---|---|---|---|
| PoseBench | Benchmark Framework | Evaluates apo-to-holo & multi-ligand docking | Standardized performance comparison across diverse scenarios [54] |
| BCAPIN | Specialized Dataset | Protein-carbohydrate complex evaluation | Assesses performance on sugar interactions [52] |
| DockQC | Evaluation Metric | Quality assessment for docking predictions | Standardized scoring for complex quality [52] |
| PLIF-WM | Specificity Metric | Measures chemical interaction accuracy | Quantifies recapitulation of native interactions [54] |
| PoseBusters | Validation Suite | Checks chemical validity of predicted structures | Identifies steric clashes and chemical irregularities [52] |
| AlphaFlow | Ensemble Generation | Creates alternative conformations | Tests robustness across conformational diversity [56] |
| MD Simulations | Structure Refinement | Molecular dynamics for model relaxation | Improves model quality through structural refinement [56] |
The comparative analysis reveals a nuanced landscape where each method exhibits distinct strengths and limitations. AF3 generally achieves high structural accuracy but demonstrates concerning overconfidence and high MSA dependence. Chai-1 shows impressive generalization with lower MSA reliance, while Boltz-1 strikes a balance between accuracy and computational efficiency. RFAA provides well-calibrated confidence scores but lags in overall accuracy, and DiffDock offers computational efficiency but lower performance on complex targets.
For researchers selecting methods for specific applications, we recommend:
- For high-accuracy prediction of standard complexes: AF3 or Chai-1 when computational resources permit and MSAs are available.
- For targets with limited evolutionary information: Chai-1 in single-sequence mode provides the most robust performance.
- For large-scale screening: DiffDock or Boltz-1 offer favorable trade-offs between accuracy and computational requirements.
- For carbohydrate-binding proteins: All methods show limitations with longer polymers, requiring cautious interpretation.
- For critical applications: Employ adversarial testing or ensemble methods to validate physical realism, particularly when exploring novel binding sites.
This comparative guide provides a foundation for method selection while highlighting the need for continued development to improve physical realism, generalization to novel targets, and performance on multi-ligand complexes. As the field evolves, integration of physical principles with data-driven approaches promises to address current limitations and further enhance the utility of these powerful tools for drug discovery and structural biology.
The advent of deep learning systems like AlphaFold 3 (AF3) has revolutionized biomolecular structure prediction, achieving unprecedented accuracy across diverse molecular types including proteins, nucleic acids, and ligands [3]. However, a critical question emerges regarding the relationship between a model's training data and its predictive performance: to what extent does AF3's accuracy depend on encountering similar structures during training? This review examines the correlation between training data similarity and prediction accuracy for AF3, specifically contrasting its performance with traditional molecular docking methods in pose prediction tasks. Understanding this relationship is essential for researchers relying on these tools for drug discovery, where accurately modeling novel compounds and targets is paramount.
Independent benchmarking studies reveal a nuanced picture of AF3's capabilities. While AF3 establishes new standards for blind prediction accuracy, its performance exhibits notable dependencies on structural similarity to its training corpus [4] [5] [37]. Concurrently, enhanced traditional docking pipelines and emerging AF3 alternatives demonstrate complementary strengths, particularly for drug-like molecules less represented in biological databases. This analysis synthesizes evidence from multiple rigorous evaluations to provide researchers with a practical framework for selecting and applying these tools based on their specific target characteristics.
Performance Comparison: AlphaFold 3 vs. Molecular Docking
Independent benchmarking reveals that AF3's performance advantage is context-dependent. On the PoseBusters benchmark, AF3 achieves a 76% success rate for ligand docking when no protein structural information is provided (true blind docking), significantly outperforming standard AutoDock Vina (approximately 41% success rate) [4] [5]. However, when enhanced with simple improvements—conformational ensembles and neural network rescoring via Gnina—traditional docking reaches 65.3% success, surpassing blind AF3 and approaching pocket-informed AF3 (72.4%) [4].
Table 1: Overall Ligand Docking Success Rates (RMSD < 2Å & PB-Valid) on PoseBusters Benchmark
| Method | Category | Input Requirements | Success Rate |
|---|---|---|---|
| AlphaFold 3 (blind) | Deep Learning | Sequence + SMILES | 76% [5] |
| AlphaFold 3 (pocket-informed) | Deep Learning | Sequence + SMILES + Pocket Residues | 72.4% [4] |
| AutoDock Vina (baseline) | Traditional Docking | Protein Structure + SMILES | ~41% [4] |
| Enhanced Traditional (Gnina + ensembles) | Traditional Docking | Protein Structure + SMILES | 65.3% [4] |
| SurfDock | Generative Diffusion | Protein Structure + SMILES | 39.3% [37] |
For antibody-antigen complexes, AF3 achieves a 10.2% high-accuracy docking success rate (DockQ ≥0.8) with single-seed sampling, substantially improving over AF2-Multimer's 2.4% but still failing in 65% of cases [7]. With massive sampling (1,000 seeds), AF3's success rate reaches 60%, highlighting the critical role of sampling intensity for challenging targets [7].
Impact of Training Data Similarity on Performance
The most compelling evidence for the memorization question comes from performance stratification based on molecular similarity to training data. AF3 demonstrates exceptional performance for "common natural ligands" (molecules appearing frequently in the PDB), but this advantage diminishes for less common, more drug-like compounds [4].
Table 2: Performance Stratification by Ligand Type on PoseBusters Benchmark
| Ligand Category | AF3 Success Rate | Enhanced Traditional Docking Success Rate | Performance Gap |
|---|---|---|---|
| Common Natural Ligands (n=50) | Highest Performance | Struggles | AF3 Superior |
| Other Ligands | Reduced Performance | 8.5% higher than AF3 | Traditional Superior |
| Halogen-Containing Ligands (n=69) | Unspecified | 84.1% | Traditional Superior |
FoldBench assessments confirm that AF3's ligand docking accuracy diminishes as ligand similarity to the training set decreases [5]. This pattern is particularly pronounced for "unseen ligands" (Tanimoto similarity <0.5 to training set ligands bound to homologous proteins), where AF3 achieves a 64.3% success rate—slightly below its overall performance [5].
For protein complexes, DeepSCFold demonstrates the value of incorporating structural complementarity information beyond sequence co-evolution, achieving 11.6% and 10.3% improvements in TM-score over AF-Multimer and AF3 respectively on CASP15 targets, and 12.4% improvement in antibody-antigen interface prediction over AF3 [57]. This suggests AF3's architectural advantages alone cannot fully compensate for lacking relevant interaction signals in its training data.
Experimental Protocols and Methodologies
Standardized Benchmarking Frameworks
Rigorous evaluation of molecular docking methods employs several established benchmark datasets and validation protocols:
PoseBusters Benchmark: Consists of 428 protein-ligand structures released to the PDB in 2021 or later, after AF3's training cutoff of September 2021 [3] [4]. The benchmark validates both geometric accuracy (ligand RMSD <2Å) and physical plausibility through the PoseBusters Python package, which checks for stereochemical violations, protein-ligand clashes, and other physicochemical constraints [4] [37].
DockQ Scoring for Protein Complexes: For antibody-antigen and other protein-protein complexes, the DockQ metric integrates interface residue contacts, ligand RMSD, and interface RMSD into a single score that correlates with CAPRI evaluation categories (incorrect, acceptable, medium, high accuracy) [7]. A DockQ score ≥0.8 corresponds to "high-accuracy" predictions [7].
FoldBench: A comprehensive low-homology benchmark that rigorously evaluates all-atom predictors by removing targets with high sequence or structural similarity to training set entries [5]. This is particularly valuable for assessing generalization to novel targets.
Method-Specific Implementation Details
AlphaFold 3 Protocol: AF3 requires only protein sequences and ligand SMILES strings as inputs for blind prediction [3]. The model employs a diffusion-based architecture that starts with noisy atomic coordinates and iteratively refines them toward the final structure [3] [24]. For optimal performance, especially on immune system proteins, multiple seeds must be sampled—the AF3 paper reported antibody docking success rates using 1,000 seeds [7] [5].
Enhanced Traditional Docking Baseline: The strong traditional docking baseline implements two key modifications to standard Vina: (1) conformational ensembles of ligands generated with RDKit to ensure diverse starting states, and (2) Gnina rescoring of output poses using a convolutional neural network trained to distinguish near-native poses [4]. This approach maintains the same training data cutoff (2017) as AF3 evaluations for fair comparison [4].
DeepSCFold Methodology: This alternative approach constructs paired multiple sequence alignments (pMSAs) by integrating sequence-based predictions of protein-protein structural similarity (pSS-score) and interaction probability (pIA-score), then uses these enhanced pMSAs with AlphaFold-Multimer for structure prediction [57]. This method specifically addresses limitations in co-evolutionary signal detection for challenging targets like antibody-antigen complexes [57].
Diagram 1: Comparative experimental workflows for AF3 and traditional docking methods, highlighting their distinct input requirements and shared evaluation frameworks.
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Computational Tools for Biomolecular Docking Research
| Tool/Resource | Type | Primary Function | Application Context |
|---|---|---|---|
| PoseBusters Python Package | Validation Tool | Validates physical plausibility and geometric accuracy of docked poses | Standardized benchmarking across methods [4] [37] |
| RDKit | Cheminformatics Toolkit | Generates ligand conformational ensembles | Enhanced sampling for traditional docking [4] |
| Gnina | Deep Learning Scorer | Rescores docked poses using convolutional neural networks | Improving pose selection in traditional docking [4] |
| DockQ | Assessment Metric | Quantifies protein complex prediction quality using CAPRI criteria | Standardized evaluation of protein-protein docking [7] |
| ABCFold | Execution Framework | Simplifies running AF3, Boltz-1, and Chai-1 with unified inputs | Comparative analysis of AF3-like models [5] |
| AlphaBridge | Analysis Tool | Post-processes and visualizes interaction interfaces in complexes | Interpreting AF3 prediction results [5] |
| DeepSCFold | Prediction Pipeline | Constructs paired MSAs using structural complementarity predictions | Handling targets lacking clear co-evolution signals [57] |
Discussion and Research Implications
The relationship between training data similarity and prediction accuracy follows distinctly different patterns for AF3 versus traditional docking methods. AF3 demonstrates superior generalization for biomolecules well-represented in its training data (common natural ligands, standard protein folds), but exhibits declining performance for novel scaffolds and drug-like compounds containing halogens or other unusual moieties [4] [5]. This pattern suggests that while AF3 has learned fundamental principles of molecular interaction, its predictive accuracy remains partially contingent on pattern recognition from similar training examples.
For traditional docking methods, performance depends more on structural complementarity and physicochemical constraints than training data similarity, making them more consistent across diverse molecule types [4] [37]. However, they require high-quality protein structures as input and struggle with binding-induced conformational changes that AF3 can potentially model through its integrated structure prediction [14] [8].
These findings suggest a synergistic approach for practical drug discovery:
- Use AF3 for truly novel targets without experimental structures or when binding sites are unknown
- Apply enhanced traditional docking when experimental structures exist and dealing with drug-like molecules
- Leverage AF3 for initial screening of binding possibilities, followed by traditional docking refinement for lead optimization
Diagram 2: Decision framework for selecting pose prediction methods based on target characteristics and available information.
Emerging AF3 alternatives like Boltz-1, Chai-1, and HelixFold-3 show promising results, with Chai-1 achieving 77% ligand RMSD success comparable to AF3, while incorporating additional features like residue-level embeddings from protein language models and trainable constraint features [5]. However, FoldBench assessments confirm AF3 maintains an approximately 10% advantage over these alternatives in protein-ligand interactions [5].
For antibody-antigen docking—a particularly challenging case—all current methods show significant limitations, with AF3 failing in 65% of cases with single-seed sampling [7]. This highlights the need for continued methodological improvements, particularly for flexible binding interfaces.
The "memorization question" reveals a nuanced relationship between training data similarity and prediction accuracy for AlphaFold 3. While AF3 represents a monumental advance in blind biomolecular structure prediction, its performance remains partially correlated with similarity to training examples, particularly for small molecule ligands. Traditional docking methods, when enhanced with modern sampling and scoring techniques, maintain competitive performance—especially for drug-like molecules less represented in AF3's training data.
These findings support a pragmatic, tool-agnostic approach to molecular docking in research and drug discovery. Rather than wholesale replacement of traditional methods, AF3 expands the computational toolbox, offering particular value for targets lacking experimental structural information. As the field evolves, the integration of AF3's holistic modeling with traditional docking's physicochemical foundations promises to advance computational structural biology most effectively. Researchers should select tools based on their specific target characteristics, using the decision framework provided to optimize prediction success.
Conclusion
AlphaFold 3 represents a monumental leap in predicting the structural landscape of biomolecular complexes, often outperforming specialized docking tools in initial pose prediction, particularly for well-represented systems. However, critical evaluations reveal its limitations in physical understanding, generalization, and handling flexibility, indicating it has not fully superseded traditional methods. The future lies not in choosing one tool over the other, but in developing synergistic workflows. These should leverage AlphaFold 3's powerful hypothesis-generation capabilities and integrate its outputs with physics-based docking, molecular dynamics simulations, and experimental validation. For researchers in biomedicine and drug discovery, a nuanced, critical, and integrated application of these technologies will be paramount for translating structural predictions into functional insights and successful therapeutic candidates.
References
- 1. Beyond rigid docking: deep learning approaches for fully ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12406700/]
- 2. FeatureDock for protein-ligand docking guided by ... [https://www.nature.com/articles/s44386-025-00005-6]
- 3. Accurate structure prediction of biomolecular interactions ... [https://www.nature.com/articles/s41586-024-07487-w]
- 4. Approaching AlphaFold 3 docking accuracy in 100 lines of ... [https://www.inductive.bio/blog/strong-baseline-for-alphafold-3-docking]
- 5. How have AlphaFold 3's predictions been validated? [https://www.ebi.ac.uk/training/online/courses/alphafold/alphafold-3-and-alphafold-server/introducing-alphafold-3/how-have-alphafold-3s-predictions-been-validated/]
- 6. Investigating whether deep learning models for co-folding ... [https://www.nature.com/articles/s41467-025-63947-5]
- 7. What does AlphaFold3 learn about antibody and nanobody ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12360200/]
- 8. AlphaFold3: An Overview of Applications and Performance ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12027460/]
- 9. AlphaFold3 and its improvements in comparison to ... [https://medium.com/@falk_hoffmann/alphafold3-and-its-improvements-in-comparison-to-alphafold2-96815ffbb044]
- 10. AlphaFold 3 Predicts Everything Now, Not Just Proteins ... [https://medium.com/@cognidownunder/alphafold-changed-biology-forever-when-it-solved-protein-folding-78bb8768483a]
- 11. Before and after AlphaFold2: An overview of protein structure ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10011655/]
- 12. Triangle Multiplication is All You Need for Biomolecular ... [https://arxiv.org/html/2510.18870v1]
- 13. AlphaFold 3 vs AlphaFold 2/AlphaFold2-Multimer [https://www.linkedin.com/pulse/alphafold-3-vs-2alphafold2-multimer-summary-genophore-inc-vuqqe]
- 14. AlphaFold3 prediction of protein-protein complex: Is it ... [https://www.sciencedirect.com/science/article/pii/S2001037025004180]
- 15. Improving peptide-protein docking with AlphaFold-Multimer ... [https://www.frontiersin.org/journals/bioinformatics/articles/10.3389/fbinf.2022.959160/full]
- 16. A workflow to create a high-quality protein–ligand binding ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11967698/]
- 17. Leak Proof PDBBind: A Reorganized Dataset of Protein- ... [https://arxiv.org/html/2308.09639v2]
- 18. Augmented BindingNet dataset for enhanced ligand ... [https://www.nature.com/articles/s44386-024-00003-0]
- 19. AlphaFold 3: an unprecedent opportunity for fundamental ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12342994/]
- 20. An Interpretable Deep Learning and Molecular Docking ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12388385/]
- 21. Assessing interaction recovery of predicted protein-ligand poses [https://pmc.ncbi.nlm.nih.gov/articles/PMC12090448/]
- 22. and Polyfluoroalkyl Substances Docking in Protein Structures [https://pmc.ncbi.nlm.nih.gov/articles/PMC12424161/]
- 23. Benchmarking Sets for Molecular Docking - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC3383317/]
- 24. Review of AlphaFold 3: Transformative Advances in Drug ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11292590/]
- 25. Reliable protein-protein docking with AlphaFold, Rosetta, and ... [https://pubmed.ncbi.nlm.nih.gov/40424178/]
- 26. A Comparative Study of AlphaFold2Multimer vs AlphaFold3 [https://pubmed.ncbi.nlm.nih.gov/39927847/]
- 27. Linear Antibody Epitope Prediction using AlphaFold2 [https://elifesciences.org/reviewed-preprints/98369v1]
- 28. Linear Antibody Epitope Prediction using AlphaFold2 - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC11042291/]
- 29. How good are AlphaFold models for docking-based virtual ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9852548/]
- 30. TEMPL: A Template-Based Protein–Ligand Pose Prediction ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12570141/]
- 31. Data-driven Priors to Improve Docking for Pose Prediction [https://chemrxiv.org/engage/chemrxiv/article-details/68a380bf23be8e43d6c26fd2]
- 32. BRANEart: Identify Stability Strength and Weakness ... [https://www.frontiersin.org/journals/bioinformatics/articles/10.3389/fbinf.2021.742843/full]
- 33. New AI Reportedly Surpasses AlphaFold 3 in Drug-Protein ... [https://www.biopharmatrend.com/news/genesis-and-nvidia-claim-pearl-surpasses-alphafold-3-in-drug-protein-structure-prediction-1420/]
- 34. Atomically accurate de novo design of antibodies with ... [https://www.nature.com/articles/s41586-025-09721-5]
- 35. AlphaFold and Glycosylation: Why Experimental Data ... [https://aspariaglycomics.com/blog/alphafold-and-glycosylation/]
- 36. Unlocking the potential of in silico approach in designing ... [https://www.frontiersin.org/journals/bioinformatics/articles/10.3389/fbinf.2025.1533983/full]
- 37. Decoding the limits of deep learning in molecular docking for ... [https://pubs.rsc.org/en/content/articlehtml/2025/sc/d5sc05395a]
- 38. Molecular docking as a tool for the discovery ... [https://www.nature.com/articles/s41598-023-40160-2]
- 39. pLDDT: Understanding local confidence | AlphaFold [https://www.ebi.ac.uk/training/online/courses/alphafold/inputs-and-outputs/evaluating-alphafolds-predicted-structures-using-confidence-scores/plddt-understanding-local-confidence/]
- 40. Highly accurate protein structure prediction for the human ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8387240/]
- 41. Analysing protein complexes in plant science: insights and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12098255/]
- 42. Ligand B‐Factor Index: A Metric for Prioritizing Protein‐Ligand ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12423484/]
- 43. A comprehensive survey of scoring functions for protein ... [https://bmcbioinformatics.biomedcentral.com/articles/10.1186/s12859-024-05991-4]
- 44. Assessment of Pharmaceutical Protein-Ligand Pose and ... [https://pubmed.ncbi.nlm.nih.gov/41045049/]
- 45. PRosettaC outperforms AlphaFold3 for modeling PROTAC ... [https://www.nature.com/articles/s41598-025-21502-8]
- 46. Docking guidance with experimental ligand structural density ... [https://pubmed.ncbi.nlm.nih.gov/39998966/]
- 47. 2025 Guide to AI-Driven Protein Design & Machine Learning ... [https://nanohelix.ai/blog/ai-protein-design-guide-2025]
- 48. No dance, no partner! A tale of receptor flexibility in ... [https://www.sciencedirect.com/science/article/abs/pii/S0065774322000069]
- 49. Flexible ligand docking to multiple receptor conformations [https://pmc.ncbi.nlm.nih.gov/articles/PMC2396190/]
- 50. AlphaFold 3 predicts the structure and interactions of all ... [https://www.isomorphiclabs.com/articles/alphafold-3-predicts-the-structure-and-interactions-of-all-of-lifes-molecules]
- 51. A Folding-Docking-Affinity framework for protein-ligand ... [https://www.nature.com/articles/s42004-025-01506-1]
- 52. Evaluation of De Novo Deep Learning Models on the Protein ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12424684/]
- 53. Benchmarking protein-carbohydrate complex prediction ... [https://www.linkedin.com/posts/karen-elliot-khar-23652a110_evaluation-of-de-novo-deep-learning-models-activity-7371250451553386496-tFd4]
- 54. Assessing the potential of deep learning for protein-ligand ... [https://arxiv.org/html/2405.14108v6]
- 55. Investigating whether deep learning models for co-folding ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12501370/]
- 56. Evaluating ligand docking methods for drugging protein ... [https://jcheminf.biomedcentral.com/articles/10.1186/s13321-025-01067-4]
- 57. High-accuracy protein complex structure modeling based ... [https://www.nature.com/articles/s41467-025-65090-7]